|
|
Главная |
|
|
Пишите |
|
Вторичноротые (лат. Deuterostomia) — подраздел (иногда надтип) билатеральных животных. К ним относятся в том числе и наиболее прогрессивные животные — позвоночные (подтип хордовых).
|
|
|
Основные особенности вторичноротых:
|
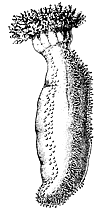
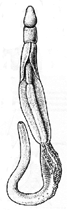
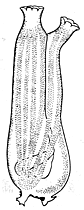
Еще один характерный для вторичноротых признак — жаберные щели. Так называются парные отверстия, которыми передний отдел кишечника открывается наружу. Функция жаберных щелей — или дыхательная (в их стенках идет газообмен), или питательная (служат для отфильтровывания из воды мелких пищевых частиц), или обе сразу. Жаберные щели есть абсолютно у всех хордовых, хотя у наземных позвоночных — только на зародышевых стадиях. Есть жаберные щели и у полухордовых. У червеобразных представителей этого типа (кишечнодышащие) они развиты очень хорошо, у сидячих (крыложаберные) могут исчезать, но это наверняка вторичное состояние. Наконец, у современных иглокожих жаберных щелей нет [а чем они дышат?], но существуют палеонтологические данные, показывающие, что у их древних родичей таковые были. Таким образом, жаберные щели — это общая черта всех основных групп вторичноротых. Вероятно, они уже были у общего предка этой эволюционной ветви.
По-видимому, уже в самом начале эволюции вторичноротых в их теле сложилась система протока воды, входившей через рот и выходившей через жаберные щели, попутно отдавая пищевые частицы.
Можно предложить для объяснения происхождения жаберного аппарата следующий внутренне непротиворечивый эволюционный сценарий:
Разделы страницы о вторичноротых животных:
Также читайте о происхождении и эволюции хордовых животных и о самых примитивных хордовых - ланцетниках.
|
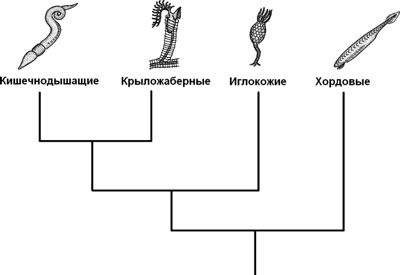
Раньше считалось, что в группу вторичноротых входят 3 типа животных: Echinodermata (иглокожие), Hemichordata (полухордовые) и Chordata (хордовые). Представители первых двух типов относятся к беспозвоночным, а к хордовым причисляют как беспозвоночных (головохордовые и оболочники), так и позвоночных (подтип Vertebrata), изучением которых занимается особый раздел зоологии. Теперь в объединение вторичноротых включают также щетинкочелюстных и погонофор, а полухордовых делят на кишечнодышащих и крыложаберных. |
Вторичноротые животные (дейтеростомы)
/ | |
/ / \ / \
/ / \ / \
Щетинкочелюстные Полухордовые Иглокожие Погонофоры Хордовые
|
|-перистожаберные (крыложаберные)
|
|-кишечнодышащие
|
|-граптолиты (ископаемые)
|
Происхождение вторичноротых неясно. Возможно они произошли от радиальных (кишечнополостных) животных независимо от первичноротых. По другим гипотезам, предками вторичноротых были представители одного из примитивных типов первичноротых, объединяемых в группу низших червей. Последние данные молекулярных исследований говорят в пользу первой теории. У вторичноротых в период зародышевого развития на месте первичного рта (бластопора) образуется анальное отверстие, а собственно рот независимо появляется в передней части тела. Есть вторичная полость тела (целом). Все билатерально-симметричные организмы делятся на две группы - первичноротые и вторичноротые. Названия "первичноротые" (Protostomia) и "вторичноротые" (Deuterostomia) произошли от способа развития ротового отверстия в эмбриогенезе. Так, у первичноротых животных бластопор (возникающее в эмбриональном развитии отверстие первичного кишечника) частично или полностью переходит в ротовое отверстие. У вторичноротых (например, у иглокожих) бластопор становится анальным отверстием, а рот прорывается заново на переднем конце личинки. В других случаях бластопор закрывается, а рот и анус открываются заново. Помимо судьбы рта, первичноротые и вторичноротые отличаются другими эмбриологическими особенностями. (1) Во-первых, это характер дробления оплодотворенного яйца: у первичноротых оно спиральное, а у вторичноротых - радиальное. (2) Другое отличие состоит в способе закладке целома (вторичной полости тела): у первичноротых стенки вторичной полости тела происходят от двух клеток, а у вторичноротых за счет выпячивания карманов эмбрионального кишечника. (3) Третьим важнейшим различием между первично- и вторичноротыми является судьба зачатка первичного мозга: у первичноротых он дает начало мозгу взрослых форм, а у вторичноротых всегда редуцируется, так что новый нервный центр возникает в другом месте заново. Вторичноротые - это вторичномозговые животные. |
Общий предок вторичноротых животных, по-видимому, вел донный образ жизни, ползая на брюшной стороне с помощью ресничной полоски, подостланной вентральным нервным тяжем. Он имел три пары целомических мешков и щупальцевый аппарат, с помощью которого собирал мелкие пищевые частицы. В дальнейшем могло произойти обособление самого заднего отдела тела, с помощью которого предки вторичноротых временно зарывались в верхние слои грунта. В эволюции вторичноротых этому отделу принадлежит очень важное место.
Очевидно, у него уже были особые отверстия, соединяющие полость глотки с внешней средой: жаберные поры. Он был фильтратором и питался бактериями и органической взвесью, которая попадала в глотку с водой через рот и осаждалась на слизистом эпителии, а вода выходила наружу через жаберные поры. Именно так и сегодня живут примитивные вторичноротые (кишечнодышащие, ланцетник, личинки миног), и такой же способ питания предполагается для древнейших ископаемых вторичноротых, таких как юннанозои и пикайя.

Ксенотурбелла (лаь. Xenoturbella) — род морских червеобразных животных до 4 см в длину, выделяемый в отдельный тип Xenoturbellida. найдены у побережья Швеции (на глубине 60—100 м во фьордах), Шотландии и Исландии. Известно 6 видов. Первые экземпляры Xenoturbella были обнаружены в шведских водах Балтики в 1915 году биологом Сикстеном Боком (1884—1946), но описаны только в 1949 году Эйнаром Вестбладом под названием Xenoturbella bocki. Последний отнес их к Acoelomorpha, которые были тогда в составе плоских червей. Известны также по имени "лиловый носок", потому что похожи именно на брошенный на пол носок. Существа эти настолько необычны, что в течение 60 лет ученые не могли определить, к какому именно виду или роду они принадлежат. Классификация и сейчас затруднительна, но предварительно их считают самыми примитивными вторичноротыми. Из-за того, что они питаются моллюсками, а в личиночной стадии паразитируют на них, их тоже сначала считали моллюсками. Сквозной пищеварительный тракт отсутствует - имеется только мешкообразная кишка без заднего прохода, образующая единственную полость тела. Чепм кушает, тем и выделяет (как гидра). Причем, ротовое отверстие довольно маленькое. Ни зубов ни сосательного хоботка нет. У них также нет централизованного мозга или нервного узла. Нервная система, расположена под эпидермисом, имеет диффузный характер. Имеется орган равновесия - статоцист. Половой системы тоже нет, но обнаружены гаметы - репродуктивные клетки, имеющие гаплоидный набор хромосом. |

Исследуя породы необычных сланцев в провинции Шаньси, которые сформировались во время "кембрийского взрыва" (около 520-510 миллионов лет назад), китайские и американские палеонтологи нашли останки необычного животного — небольшой овальной структуры длиной в один миллиметр, похожей на мешок с "зубастыми" краями. Оно было названо Saccorhytus coronarius, что переводится с латыни как "сморщенный мешок с короной". Эти организмы, как предполагают ученые, обладали неким подобием жабр, следы которых можно заметить в виде конических структур на его коже. Через них "мешок" выпускал воду, которую проглатывал вместе с пищей. Впоследствии эти конусы могли превратиться в жаберные дуги, а затем — в челюсти и нос первых рыб. |
Простейшие | Пластинчатые | Губки | Гидры | Линяющие | Спиральные | Плоские | Щупальцевые | Вторичноротые
Ключевые слова для поиска сведений о вторичноротовых животных:
На русском языке: вторичноротовые животные, вторичноротые беспозвоночные,
вторичный рот, прогрессивные позвоночные, подтип хордовых, иглокожие, полухордовые, щетинкочелюстные,
радиальное дробление оплодотворенного яйца;
На английском языке: Deuterostomia.
|
|
|
|
|
|