|
|
Главная |
|
|
Пишите |
|

Все, что можно основательно поколебать - необходимо поколебать, и если оно не устоит - так тому и быть. (Будда) [принцип эволюции?] Глобальные биотические кризисы - это относительно длительные события, занимающие 10-15 млн. лет. Дестабилизирующий биоту толчок приводит к вымиранию специализированных господствующих видов, происходит упрощение структуры сообщества и сокращения разнообразия. При последующей стабилизации обстановки восстанавливаются прежние тенденции, быстрый рост разнообразия создает впечатление "взрывной эволюции" [Красилов и др., 1985]. Глобальные биотические кризисы представляют собой критические моменты существенной перестройки всей биосферы. Важнейшей составной частью биотических кризисов являются лежащие в их основе массовые вымирания (МВ), благодаря которым освобождаются ранее занятые экологические ниши и происходит "омоложение" сообществ различного уровня, что обеспечивает ускоренную эволюцию переживших эти события групп организмов [Алексеев, 1989]. |
Разделы страницы о биотических кризисах, их закономерностях и причинах:
О космических причинах катаклизмов на Земле читайте в разделе о Солнечной системе в целом (история падений метеоритов, солнечная активность...). А сами эти процессы циклически зависят от вращения нашей планетной системы вокруг центра Галактики, о чем читайте в авторской статье о галактических циклах. От этих же факторов зависят и геологические циклы.
Выживает не самый сильный и не самый умный, а тот, кто лучше всех приспосабливается к изменениям. (Чарльз Дарвин)
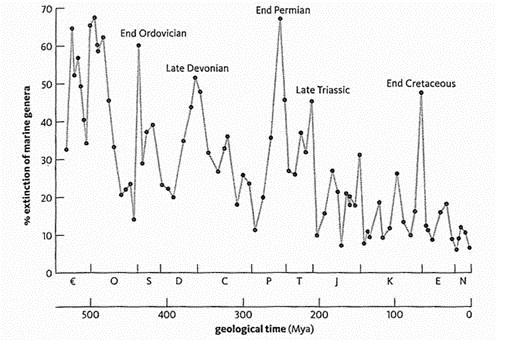
Биотические кризисы и сопровождавшие их МВ значительно различались по масштабам. Среди них отчетливо выделяются 4-5 наиболее крупных событий - "великие массовые вымирания" и около 15-ти событий значительно меньшей амплитуды и продолжительности. Это так называемые "малые вымирания". Всего же можно насчитать до 29 событий МВ. К великим массовым вымираниям, которые однозначно признаются всеми исследователями, можно отнести следующие четыре: мел-палеогеновое (2), триасовое-юрское (3), пермско-триасовое (4) и ордовикско-силурийское (6). Девонская катастрофа тоже была весьма крупной, но охватывала только один из двух суперматериков (Гондвану), поэтому, видимо, ее и не включают в число "великих вымираний". Тем не менее, триасовое вымирание, хоть и было "великим", но охватывало только морские виды, поэтому не являлось всеобщим, хоть и охватывало все климатические широты Земного шара. Отсюда следует, что истинно глобальными биокатастрофами фанерозоя были ордовикская, пермская, меловая, интервалы между которыми, на минуточку, примерно равны Галактическому году, о чем подробнее ниже. |
.png)
Самые древние (протерозойские и архейские) биокризисы пока смутно выявляются только косвенными способами. В 1994-1996 г.г. М.А.Семихатов и М.Е.Раабен опубликовали серию статей, в которых подвергли анализу мировые данные по строматолитам архея и протерозоя (3,8-0,545 млрд. лет). Установлена довольно сложная история цианобактериальных экосистем – на фоне растущего обилия и разнообразия строматолитов в течение архея и протерозоя было три спада. Последний из них начался около 850 млн. лет назад и оказался фатальным, т.к. совпал с началом длительной канадской ледниковой эры. По мнению авторов, именно климатический фактор был ведущим в элиминации строматолитовых сообществ в качестве доминант относительно мелководных бассейнов. Эти данные были также графически представлены Е. М. Хабаровым [Хабаров 2011, с. 1459]. |
|
|
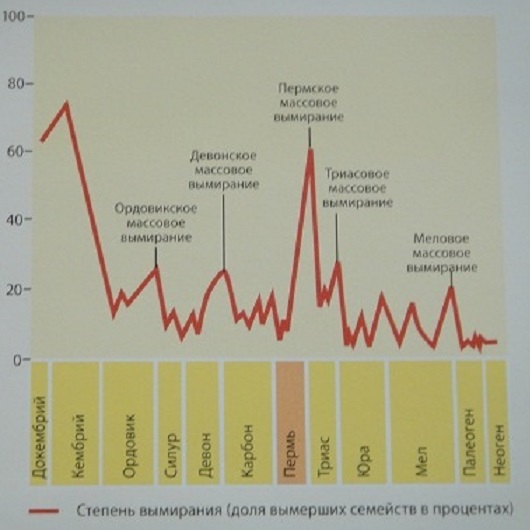
| Степень вымирания (доля вымерших семейств в %) в фанерозое |
Массовые вымирания в фанерозое (последние 0,5 млрд. лет) Источник: Википедия. |
Всего пока зафиксировано 10 биологических катастроф, большинство из которых неузнаваемо изменили облик биосферы всей планеты, либо отдельных её оболочек или отдельных материков. [Например, последняя, плейстоценовая, погубила мегафауну Евразии, но не затронула, например, Африку.] При этом гибло от 1/4 до 3/4 и более биологических видов. Недавно появились данные о плиоценовом биокризисе - промежуточным между "мамонтовым" (плейстоценовым) и "динозавровым" (меловым). Биоатастрофы ниже перечислены от поздних к ранним.
Фанерозойские (за последние полмиллиарда лет):
Докембрийские:
"Бог следит внимательно за теми процессами, которые происходят на Земле, и пока эти процессы развиваются в соответствии с задуманным сценарием он остаётся сторонним наблюдателем. Но стоит эксперименту принять нежелательный поворот, как следует оперативное вмешательство." (Лимарев)
Периодичность МВ принадлежит к числу наиболее дискуссионных вопросов, так как для количественного анализа временных рядов требуются точные исходные возрастные датировки, которые иногда крайне трудно получить даже для одной границы. Несмотря на противоречивые результаты статистических исследований, имеются все основания считать, что глобальные биотические кризисы, частью которых являются МВ, могли быть периодическими вне зависимости от их природы, что вытекает из периодического характера большинства земных и космических процессов. Высказывалось также мнение, что наряду с периодическими малыми МВ отмечаются апериодические великие МВ [Алексеев, 1989].
Особенно активно эта проблема стала дискутироваться после публикации статьи Д.Раупа и Дж. Сепкоски [Raup, Sepkoski, 1984], в которой постулировалось существование отчетливо выраженной периодичности МВ в мезо-кайнозое с интервалом около 26 млн.лет. Различные исследователи тут же выступили с критикой этого положения, опираясь на неправильный методологический подход авторов [Алексеев, 1989, с. 40]. Указанная американскими авторами периодичность с интервалом близким к 30 млн. лет существует, но в то же время не всегда выдерживается. Так в течение фанерозоя отмечаются периоды некоторого сгущения МВ, например во второй половине девонского периода, конце перми и конце триаса.
Также имеется мнение, что залёты крупных метеоритов на Землю тоже случаются примерно раз в 30 миллионов лет. Необходимо заметить, что и осцилляции орбиты Солнечной системы относительно плоскости Галактики также идут с периодичностью около 30 млн лет.
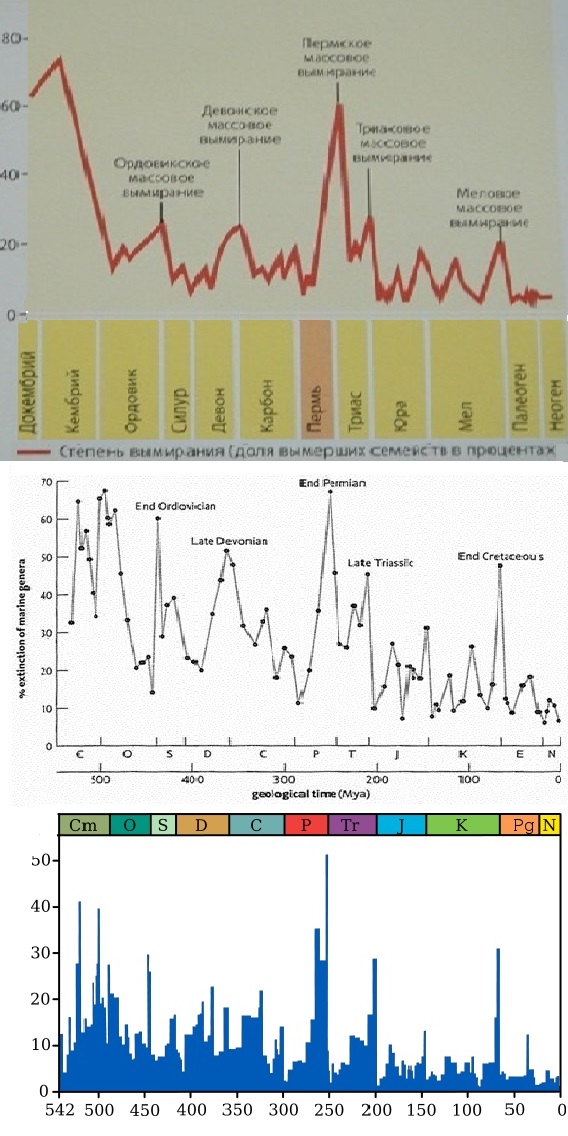
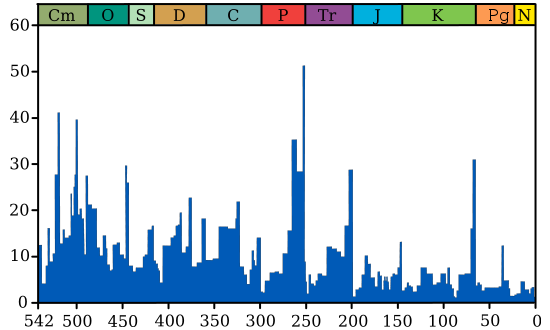
Первые 6 из перечисленных 10 глобальных биокризисов произошли за последние 500 миллионов лет (в фанерозое). Они смели с поверхности Земли от 50 до 96% флоры и фауны. Следующие 4 - за период 3,8-0,545 миллиардов лет, т.е. докембрийские. По ним меньше сведений [да и биологических катастроф в докембрии, скорее всего, было много больше]. Для 4 из представленных биоценозных суперкатастроф предподагается импактная причина. В то же время, как минимум, 6 из обсуждаемых здесь 10 планетарных биокатастроф (в т.ч. 2 докембрийские) связаны с эпохами оледенений. Однако, позднекарбонский ледниковый период (гондванское оледенение), произошедшее около 350-230 м.л.н. (максимум пришелся на 280 м.л.н.), как будто, не привело к катастрофическим событиям в биосфере Земли. [На самом деле была, пусть не глобальная, но значительная гибель биоты 270 млн. лет назад.] А самое, наверное, впечатляющее совпадение - в том, что 7 из этих 10 супербиокризисов случились во время или рядом (перед) прохождением Солнечной системы через перигалактий - галактический перигелий, что случается примерно через каждые 200 миллионов лет. С перигалактием не связаны лишь четвертичная, триасовая и девонская биокатастрофы, которые не носят общепланетарный характер: первая не проявилась в Африке, вторая касалась только гидросферы, третья была только в Гондване. На всех 3-х графиках бросается в глаза примерно одинаковая длительность между ордовикской (440-450 Ma), пермской (250 Ma) и меловой (65-70 Ma) биокатастрофами (их соединяет зеленая прямая линия): 190 и 180 миллионов лет. Такая же примерно длительность - между промежуточными пиками на этих интервалах, а именно, между девонским биокризисом (~365 Ma) и пиком около 140-145 млн. лет (их соединяет синяя прямая): интервал в 220 млн лет. Т.о. прослеживается период около 100 млн. лет (фиолетовая прямая). В то же время на 3-м графике видно, что ордовикско-пермский и пермско-меловой интервалы делится пиками на 3 части (коричневая прямая). В этом случае подциклом будет 60-70 млн лет. Видно, что наиболее крупные катаклизмы на Земле случаются примерно каждые 200 миллионов лет, что равно циклу обращения Солнечной системы вокруг центра нашей Галактики ("галактическому году"). Этот период также соответствует геотектоническому циклу Бертрана. Кроме этих глобальных биокатастроф эпизодически просходят и более локальные биокризисы, не связанные с перигалактическим фактором. |
По-видимому, биотические кризисы имели различную амплитуду и продолжительность, но последовательность событий при этом выдерживалась. Модель структуры глобального биотического кризиса была предложена еще в 1924 г. [!] Д.Н.Соболевым. Она включала в себя:

В последние годы разработке этой проблемы особое внимание уделяет О. Валлизер [Wallizer, 1986, 1995]. Всестороннему рассмотрению этой проблемы посвящены исследования в рамках международного проекта N 335 Международной программы геологической корреляции. (I) Начало кризиса совпадает с неким весьма кратковременным глобальным событием, которое дестабилизирует биосферу. В результате этого события, резко изменяющего условия обитания организмов, начинается их МВ, распространяющееся по пищевым цепям. Само вымирание занимало, как правило, довольно значительный промежуток времени (до нескольких сотен тысяч лет). Растянутость МВ вполне понятна, так как реакция сложно построенной биосферы не может быть мгновенной. Необходимое для реакции биосферы время составляло иногда сотни тысяч лет, но возможно, и значительно больше. Поэтому МВ при рассмотрении их в более крупном масштабе обычно хорошо определимы и приобретают ступенчатую структуру. (II) За ступенчатым вымиранием следует довольно длительный (от 1-2 до 5-6 млн. лет) период низкого таксономического разнообразия, когда вымирание доживающих таксонов уравновешивается появлением новых. Лицо биоты определяют древние группы, которые оказались способными пережить неблагоприятный момент резких изменений среды, однако эволюционно они, как правило, бесперспективны. (III) Наконец, весь процесс кризиса завершается бурной диверсификацией, быстрым ростом разнообразия глобальной биоты, благодаря занятию освободившихся экологических ниш. Как правило, через 5-6 млн. лет после начала диверсификации достигается, либо даже превышается разнообразие, существовавшее на предкризисной стадии. |
Модель изменения состава биотического сообщества при массовом вымирании была предложена Э.Кауффманом [Kauffman, 1994]. В ней выделено несколько групп таксонов, отличающихся по своей тактике по отношению к кризисной ситуации. В зависимости от преобладания определенных групп выделяются этапы в структуре глобального биотического кризиса:
Особая роль в структуре массового вымирания принадлежит "переживающим" и "лазерным" таксонам, дающим в последствии начало новому эволюционному этапу.
Наиболее распространенные гипотезы о причинах биологических катастроф:

В начале 80-х годов была выдвинута гипотеза о том, что массовое вымирание организмов, приуроченное к рубежу М/Д (маастрихт/даний ), было вызвано последствиями столкновения с Землей крупного космического тела, астероида, чей диаметр мог достигать 10 км [Alvarez et al., 1980]. Свидетельством импактного события послужила так называемая "иридиевая аномалия", первоначально выявленная на границе М/Д в Италии (Губбио) и Дании (Стевенс Клинт). Иридий, сидерофильный элемент из группы платиноидов, имеет крайне низкую концентрацию в земной коре - около 0,03 нг/г [Назаров и др., 1988], но весьма распространен в космическом веществе, где его содержание колеблется в пределах 500-5000 нг/г. В пограничной глине концентрация иридия чаще всего составляет от 2 до 80 нг/г, т.е. на два-три порядка выше, чем ниже и выше по разрезу. Так называемая "иридиевая аномалия" ныне установлена примерно в 150 разрезах, рассеянных по всему земному шару в океанах и на континентах. Космическая природа данной геохимической аномалии аргументируется также
Достаточно полную картину геологических и биологических последствий импактных событий дают McLaren, Goodfellow [1990]. С различной степенью обоснованности и вероятности они связывают массовые вымирания на рубежах мела-палеогена ("мезозойское" 70 Ma), эоцена-олигоцена (34 Ma) [не слышал про такой - вот в конце олигоцена был], триаса-юры (210 Ma), перми-триаса (251 Ma), девона-карбона (364 Ma), франа-фамена (375 Ma), ордовика-силура (450 Ma), докембрия-кембрия (600 Ma) с гигантскими ударными событиями. По приводимым авторами данным внеземные тела диаметром 10 и более километров сталкивались с нашей планетой в течение фанерозоя в среднем один раз в 55 млн. лет. |
Выявлена периодичность вымираний в 500 и 60 млн. лет. Другие исследования дают похожие результаты. Каждые 62 млн лет [~ 1/4 - 1/3 галацикла] появляются, переживают расцвет и погибают новые биологические виды.
Раз в 60 миллионов лет материки планеты поднимаются [прилив от массивного космического тела?], чтобы затем вновь опуститься на прежнее место. Когда бы ни произошло массовое вымирание, каждый раз в его жертвах обнаруживается повышенное содержание стронция-87. Когда происходят материковые подъемы, в море попадает много стронция-87, увеличивая его содержание по отношению к стронцию-86. [Может быть, не от ударов метеоритов, а от осколков сталкивающихся небесных тел?]
Профессор Роберт Фрей из Копенгагенского университета вместе с коллегами разработали новую методику изучения геологического прошлого планеты (журнал Nature). Для этого они проследили за состоянием атмосферы Земли за последние 3,8 миллиардов лет. А особенно — за кислородом. Увеличение концентрации кислорода в атмосфере приводит к окислению хрома в горных породах Земли. И по этим «зарубкам» исследователи и решили точно выяснить, когда на нашей планете стали жить первые организмы и какими они были.
Хронология появления кислорода известна:
То есть сегодня наш воздух состоит на 21% из кислорода и на 78% из азота. Остальное — гелий, аргон, неон, метан и углекислый газ.
Таким образом, отсчитав хронологию и количество газов в воздухе, ученые выяснили, что первые простейшие бактерии, способные к фотосинтезу, появились за 300 млн. лет до кислородной катастрофы — резкого повышения концентрации кислорода в атмосфере Земли в начале протерозоя. Эти уникальные микробы могли каким-то образом легко обходиться без жизненно важного для современных организмов газа.
Также удалось выяснить, что первоначальное накопление кислорода в атмосфере проходило постепенно и началось задолго до кислородной катастрофы, еще 2,8-2,6 млрд. лет назад — гораздо раньше, чем было принято считать. И еще один поразительный факт: через 500 млн. лет после катастрофы уровень кислорода упал до ничтожно малого, а затем вырос до современных значений. Причина этого феномена пока не ясна.
Точно было установлено, что современная концентрация кислорода в воздухе и количество растворенного газа в океанах достигли нынешнего уровня примерно 580 млн. лет назад [не стыкуется!]. И после этого началась настоящая жизнь на нашей планете. [Есть мнение, что воздух стал современным уже 1,8 миллиард лет назад!]
Если представленные данные верны, придется пересмотреть современные представления об условиях окружающей среды, сложившихся 1,9 миллиардов лет назад.
В геологической истории продолжительность отдельных пульсационных циклов и их особенности не оставались постоянными, а зависели от положения данного цикла в структуре более крупных пульсационных циклов. Наиболее важным результатом описываемых сверхглубинных процессов явились многократные небольшие изменения объема Земли и общей площади земной коры, приводившие к деформациям сжатия и растяжения, а также многократные изменения интенсивности магматизма, связанные как с неравномерным во времени волнообразным характером распространения из глубоких недр к поверхности порций глубинного тепла и вещества, так и с адиабатическими процессами в очагах магмогенерации, в частности, с декомпрессией во время экспансионных фаз геопульсаций. С одним из таких глобальных пульсационных эпизодов А.Б. Веймарн и Е.Е. Милановский [1993] связывали катастрофические события на рубеже франского и фаменского веков [девонская катастрофа?].
Роль аномально нагретого глубинного вещества, струи которого в виде "мантийных плюмажей" (плюмов) поднимаются от ядра Земли, специально рассмотрена Н.Л. Добрецовым [1994]. Он полагает, что интенсивность течений и тепломассопереноса в жидком ядре не компенсируется медленными течениями в нижней мантии. В результате внешнее ядро "перегревается", в нем повышается частота магнитных инверсий. Но, как только отрывается мантийный плюм от границы ядро-мантия, он уносит избыток тепла, внешнее ядро охлаждается, частота магнитных инверсий снижается, либо они вовсе исчезают, как это имело место в мелу и перми. Отрыв крупных плюмов (или группы плюмов) происходит периодически каждые 30 млн. лет на фоне более длительной периодичности 280-200 млн. лет. Поднимающиеся мантийные плюмы в периоды их максимальной интенсивности, сопровождаемой интенсивным мантийным магматизмом, перестраивают астеносферные течения, что вызывает периодические изменения скорости и (или) направления движения литосферных плит. Это приводит к ускорению субдукционно-акреционных процессов на конвергентных границах плит, что объясняет периодичность максимумов возрастов глаукофановых сланцев, островодужного вулканизма и оруденения.
В свою очередь, изменение рельефа в океанах и на континентах приводит к коррелирующимся процессам трансгрессий-регрессий и изменения течений в гидроатмосфере, вызывающих климатические изменения, и связанные со всем этим палеогеографические и палеоэкологические события. Огромно влияние на экологическую обстановку катастрофических вулканических извержений. Корреляция фаз реорганизации литосферных плит и короткопериодических изменений уровня Мирового океана и связанных с этим глобальных регрессий и массового вымирания организмов за последние 80 млн. лет охарактеризована в работе Е.Е. Милановского, А.М. Никишина с соавторами [1992]. Исходя из этого, Н.Л. Добрецов [1994] считает, что эндогенные процессы являются первопричиной, наиболее удовлетворительно объясняющей массовые вымирания биот и климатических кризисов.
Е.Е. Милановский [1995], рассматривая процессы, протекающие во внешнем жидком ядре и у его границы с мантией, считает, что именно они ответственны за наблюдаемые на поверхности Земли и в ее коре тектонические, магматические и другие проявления геопульсаций. Последние характеризуются чередованием преобладания в глобальном масштабе тектонических деформаций горизонтального расширения коры (фаз активизации рифтинга и спрединга) и ее сжатия (фаз складчатости). При этом из соотношения инверсий магнитного поля с фазами геопульсаций вытекает, что передача глубинных энергетических импульсов к поверхности должна была осуществляться почти мгновенно посредством изменения напряженного состояния всех оболочек Земли.
В 1982 г. вышла книга С.Г. Неручева "Уран и жизнь в истории Земли". Автор полагает, что основные особенности ураноносных черных сланцев (стратиграфическое положение, фациальная природа, исчезновение многих видов фауны и флоры) можно объяснить, исходя из того, что главная причина образования черных сланцев - периодическое заражение бассейнов седиментации (и биосферы в целом) эндогенным ураном и тяжелыми металлами токсикантами. По М.Г. Неручеву большинство черных сланцев - продукт вырожденных экосистем, приспособившихся к условиям повышенной радиации и повышенной концентрации токсичных элементов. При этом обычно стимулируется экстремальная биопродуктивность простейших водорослей и происходит резкое обеднение видового состава более высокоорганизованной фауны. "Проявление эпох уранонакопления, а одновременно фосфатонакопления и интенсивного накопления планктоногенного органического вещества, контролируется, вероятно, общим пульсационным ритмом развития Земли и осуществляется периодически в эпохи растяжения, сопровождающиеся активизацией рифтогенеза и вулканических процессов" [Неручев, 1982, стр. 197]. Для этих эпох характерен интенсивный вынос из подкоровых глубин P, U, V, Ni, Cu, Co и ряда других элементов, что вызывало соответствующую ответную реакцию биосферы. На протяжении фанерозоя С.Г. Неручев выделил 20 таких эпох и показал, что они могут быть сопоставлены с охарактеризованными к тому времени Е.Е. Милановским [1978] этапами активизации рифтогенеза. В большинстве случаев по С.Г. Неручеву ураноносные эпохи разделены во времени фазами складчатости.
А.А. Маракушев на основе детального изучения метеоритов и обширных экспериментальных исследований последовательно развивает идею о дегазации ядра Земли [1992, 1997 и др.]. По его мнению периодические усиления эндогенной активности Земли определяются сожными взаимоотношениями твердого субъядра, кристаллизующегося в ее центре и жидкого флюидного ядра, генерирующего магнитное поле. Кристаллизация никель-железного субъядра сопровождается накоплением на фронте кристаллизации совместно с водородом многих флюидных компонентов (серы, галогенов, углеводородов) и рудных металлов. Важным следствием кристаллизации является непрерывное повышение общего флюидного давления в жидком ядре. Рост давления приводит при достижении критического значения к выбросам водородных флюидов (плюмов) в мантию и земную кору, которые подвергаются флюидному плавлению с образованием в них магматических очагов. Эти импульсы дегазации ядра, искажающие нормальное развитие магнитного поля, соответствуют усилению эндогенной активности Земли. К наиболее ярким проявлениям этого процесса относится образование взрывных кольцевых структур на платформах, характеризующихся крайне высоким энергетическим уровнем взрывов, который фиксируется образованием в них алмаза и диаплектовых стекол. Эти структуры порождаются быстрым "сквозным" подъемом флюидов из жидкого ядра Земли.
Огромная потенциальная энергия сжатия, заключенная в восходящих флюидных потоках, реализуется помимо взрывов образованием структур растяжения и обновления земной коры и мантии. К ним относятся: 1) мировая рифтовая система срединноокеанических хребтов, 2) периокеанические и периконтинентальные рифты, 3) окраинные моря и другие тефрогенные структуры на континентах. Магматические формации в структурах растяжения связаны с активизацией верхней мантии восходящими флюидными потоками, обусловившими развитие в ней магматических очагов и конвективных ячеек, которые обеспечивают растяжение земной коры, ее разрушение и замещение веществом мантии.
В восходящих флюидных потоках осуществляются реакции диспропорционирования (H2 + CO = C + H2O; H2 + 3CO = 2C + CO2 + H2O и др.). При этом реализуется главная функция водорода - генерация воды, которая играет роль не только в образовании Мирового океана, но и в возникновении глубинных очагов магматизма. С понижением температуры равновесия реакций смещаются вправо и кроме воды образуется свободный углерод. Поэтому импульсы интенсивной дегазации земного ядра, при которых углеродосодержащие газы мигрируют вверх, фиксируются в осадочной оболочке Земли горизонтами черносланцевых формаций, имеющих глобальное распространение. Им же подчинен [что значит и каким образом?] комплекс стратиформных месторождений с широким спектром рудных металлов. А.А. Маракушев и др. [1997] подчеркивают, что традиционные решения проблемы генезиса рудоносных углеродсодержащих осадков акцентируют роль органического вещества. Однако, углеродсодержащие осадки обогащены таким разнообразным комплексом химических элементов, которые определенно свидетельствуют о вовлечении в их образование глубинных процессов. Это подтверждается и обнаруженными в последние годы [Томсон, Тананаева, 1994] приразломными (рвущими) черными сланцами, геохимия которых аналогична осадочным черным сланцам.
С кембрия "...впервые устанавливается временная связь черносланцевых формаций и биотических кризисов, нарушающих нормальную эволюцию животного мира..." [Маракушев, 1997; стр. 498].
Развивая взгляды А.А. Маракушева о дегазации ядра Земли, В.Л. Сывороткин в своей работе "Рифтогенез и озоновый слой" [1996] собрал и систематизировал данные по потокам природных газов (водорода, метана, гелия и др.), провел корреляцию центров дегазации планеты и наиболее устойчивых озоновых аномалий, оценил возможные термические и динамические эффекты взаимодействия эндогенных флюидов с газами гидро- и атмосферы. На основании этого им сделан целый ряд выводов, среди которых необходимо отметить следующие:
Возможно, биологические катастрофы возникают из-за антиадаптации биоты следующей фазы климато-химического колебания. Геологическая история пронизана циклами (вложенными и независимыми, экзогенными и эндогенными), а также непериодическими стихийными явлениями (обычно космическими), часть из которых может носить колебательный характер (2-фазная одиночная волна).
В зависимости от интенсивности и длительности таких процессов биотический кризис может иметь разный характер:
В двух последних случаях биокризис носит характер биокатастрофы, которая, в перспективе, весьма вредна для биосферы, так как могут потеряться не только отряды и семейства, но и классы и типы живых существ. После такого "бутылочного горлышка" следующий биокризис переживать сложнее.
Явления, обратные биокризисам по количеству, размерам и качеству, являются (авт. И.г.):
Интересно и по этим процессам выявить как причины, так и периодичность. Возможно, здесь будет связь с факторами и циклами биокатастроф.
В современной зоологии и палеонтологии мегафауной называют совокупность видов животных, чья масса тела превышает 40-45 кг. Таким образом, формально в понятие мегафауны входят существа, которых не принято считать гигантами, напр., рыжие кенгуру, а также люди. На практике, однако, под мегафауной подразумеваются виды, значительно превосходящие человека по размерам, в частности, крупнейшие сохранившиеся дикие животные Земли: слоны, жирафы, носороги, бегемоты и так далее. К мегафауне также относятся вымершие крупные птицы и млекопитающие: моа, тур и другие [мамонт, шерстистый носорог, гигантский ленивец...]. (Википедия)
Русскоязычные труды о массовых вымираниях, их возможных причинах и сопутствующих факторах [у нас, как правило, более глобальный или разнообразный подход].
В основном, англоязычные работы о глобальных вымираниях жизни - здесь чаще исследуют влияние геофизических и астрофизических факторов - космического излучения (циклического или эпизодического - от ближайших нейтронных звёзд, из центра Галактики и даже из внегалактических источников) и геомагнитного поля [т.е., бьют не по площадям, а по глазам, что имеет и плюсы и минусы].
Главная Науки о природе Биология :
Биохимия | Биогенез | Генетика | Эволюционная теория | Биокризисы | Палеонтология | Биоадаптации | Биосфера | Ультрамикробы | Вирусология | Микробиология | Ботаника | Микология | Зоология | Биопорталы | Биоцентры | Биотермины | Биокнига | Работы автора
Ключевые слова для поиска сведений о биокризисах и супербиокатастрофах:
На русском языке: биологические катастрофы, глобальные биотические кризисы, причины массового вымирания живых организмов,
биокатастрофы, биокризисы, кризисные периоды развития биоты, биоценозные суперкатастрофы;
На английском языке: biological catastrophe, biotic crisis, great extinctions.
|
|
|
|
|
|