|
|
Главная |
|
|
Пишите |
|
"...Бог следит внимательно за теми процессами, которые происходят на Земле, и пока эти процессы развиваются в соответствии с задуманным сценарием он остаётся сторонним наблюдателем. Но стоит эксперименту принять нежелательный поворот, как следует оперативное вмешательство." (Лимарев)
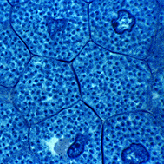
Прокариоты (Prokariota) - доядерные организмы, не обладающие типичным клеточным ядром и хромосомным аппаратом. К ним относятся бактерии, синезелёные водоросли (цианобактерии), риккетсии, микоплазмы и др. По новейшей системе органического мира, царству (или надцарству) прокариот противопостоит царство (или надцарство) эукариот [вирусы не относятся ни к одному]. Размеры прокариотных клеток в большинстве случаев колеблются от 0,5 до 3 мкм, при этом самые мелкие (микоплазмы) не превышают 0,10–0,15 мкм. Однако, гигантские клетки некоторых серобактерий достигают 100 мкм в длину, а клетки спирохет иногда вырастают до 250 мкм. Наследственный механизм представлен у прокариот нитью ДНК [или РНК], обычно кольцеобразной формы и локализованной более или менее в центральной части организма. Эта часть (нуклеоидом) не отграничена мембраной от цитоплазмы. ДНК, в отличие от эукариотов, обычно не связана с белками гистонами (не образует нуклеогистон), и регуляция работы генов осуществляется через метаболиты. |
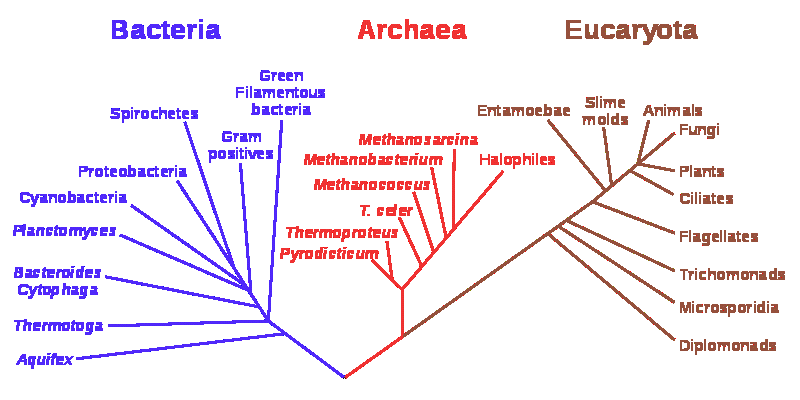
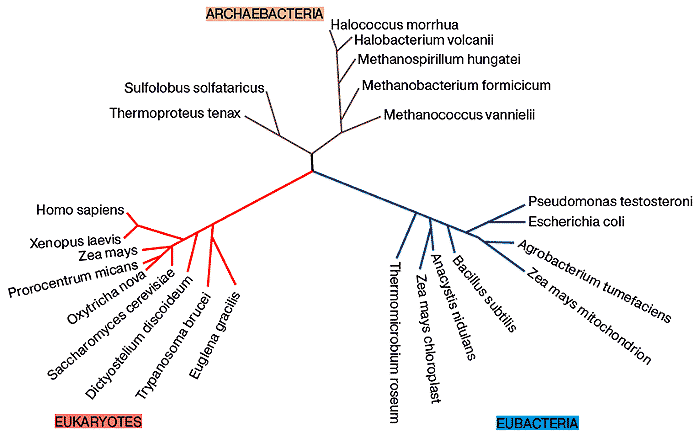
Archaebacteria и Eubacteria часто объединяются в единую группу бактерий или, иногда, монер (Moneres - греч., одинокий). Известно свыше 5 000 видов, представляющих собой маленькие клетки без ядра и органоидов.
В настоящее время описано около 10 000 видов прокариот, а настоящее их число, возможно, порядка 100 000! Многие плавающие полипептидные последовательности пока не идентифицируются, т.е., к ним еще не подобраны среды для культивации - поэтому все они скопом называются "некультивируемыми видами" (не путать с "некультивируемыми формами", т.е., анабиозными формами бактерий. (Из палеофорума - https://paleoforum.ru/?topic=1136.0)
Разделы страницы о неядерных одноклеточных микроорганизмах:
Главная черта прокариот — отсутствие ядра. Их генетический материал (генофор) представлен единственной кольцевой молекулой двухцепочечной ДНК, закреплённой на цитоплазматической мембране, одевающей клетку. У прокариот нет ядерной оболочки, эндоплазматического ретикулюма (иногда имеются впячивания поверхностной мембраны — т.н. мезосомы), митохондрий, пластид и других цитоплазматических органелл, свойственных эукариотам. У них отсутствуют и микротрубочки, поэтому они не имеют ни центриолей, ни веретена деления. Рибосомы прокариот лишены одного из типов рибосомальной РНК (так называемой 5,8S РНК) и имеют меньшую массу, чем у эукариот. Обычно масса рибосом оценивается так называемой константой седиментации (показателем скорости оседания при центрифугировании). Для рибосом прокариот она равна 70S, а для эукариот — 80S.
Прокариоты, по сравнению с эукариотами, обладают громадным разнообразием обменных процессов. Они способны к фиксации углекислоты, азота, различным вариантам брожения, окислению всевозможных неорганических субстратов (соединений серы, железа, марганца, нитритов, аммиака, водорода и др.). Среди прокариот немало фотосинтезирующих форм, прежде всего это часто встречающиеся в современной биосфере цианобактерии, которые ещё называют сине-зелёными водорослями.
Они (или родственные им организмы) были широко распространены и в далёком прошлом. Геологические постройки, созданные древними цианобактериями (вероятно, вместе с другими фотосинтезирующими прокариотами) — строматолиты, — нередко обнаруживаются в древнейших слоях земной коры, соответствующих архею и раннему протерозою.
В конце 80-х годов прошлого века в Палеонтологическом институте им. А.А. Борисяка РАН под руководством А.Ю. Розанова было создано новое направление палеонтологии – бактериальная палеонтология. Ее областью интересов являются ископаемые прокариотные микроорганизмы и их взаимоотношения с вмещающими породами, а основным методом исследований – электронная микроскопия (сканирующие электронные микроскопы с микроанализаторами).
Первым объектом бактериальной палеонтологии в ПИНе стали нижнекембрийские фосфориты Хубсугульского месторождения в Монголии, которые до наших исследований считались эталоном хемогенных фосфоритов. Уже первые полученные результаты были очень показательны. Было установлено, что микрозернистые фосфориты сложены мелкими желвачками размером десятки или первые сотни микрон, которые представляют собой фосфатизированные фрагменты цианобактериальных матов, реже онколитов.
В дальнейшем была проделана большая работа по изучению этих фосфоритов. Были просмотрены образцы, детально отобранные по всему разрезу, изучены все типы фосфоритов данного месторождения. Кроме этого начались наши совместные работы с микробиологами группы академика Г.А. Заварзина из Института микробиологии им. С.Н. Виноградского, которые помогли точно идентифицировать наши находки. В результате был издан Атлас, посвященный микроорганизмам из древних фосфоритов Хубсугула (Монголия). И эти фосфориты стали первым модельным объектом бактериальной палеонтологии. В дальнейшем было продолжено изучение фосфоритов разного возраста и из разных регионов мира. (Е.А. Жегалло)
Если группировать доядерные одноклеточные (и вирусоидные с нанобактериями, для сравнения) организмы по размерам, то градация такая:
Поскольку организмы восприимчивы к аустическим и электромагнитным (ЭМ) колебаниям, то для диапазона 0,01-3 мкм получим следующие частоты звуковых и ЭМ излучений: более 480 МГц для звука (в природе этот гиперзвук возникает при колебаниях молекул в узлах кристаллической решетки) и от ультрафиолетового света до рентгеновского излучения для ЭМИ.
Эукариоты уже будут резонировать с инфразвуком и электро-магнитными микроволнами. В целом же, получается, что вся шкала света от ультрафиолетового до инфракрасного нужна для восприятия эукариотическими организмами, т.к. ЭМИ этих частот активно воздействует на эукариотическую клетку.
Что касается бактерий, то мелкие из них резонируют с рентгеновским излучением, поэтому, возможно, в их зрительных органах (если такие есть) должны восприниматься и X-лучи. В то же время прокариоты воспринимают гиперзвук (поток фононов), длина волны которого равна среднему пробегу молекулы до ее столкновенияч с другой - а это значит, что в бактериях возможен обмен неискаженными сигналами с помощью броуновского движения.
Считается, что в очень далёком прошлом все три домена жизни – бактерии, археи и эукариоты [а микоплазмы и риккетсии разве не домены?] – сходились в одной точке, которая называется последним универсальным общим предком, или LUCA (Лука) – last universal common ancestor. Лука жил на Земле примерно 3,5–3,8 млрд лет назад, и в нём уже были запечатлены все основные черты земной жизни: его наследственная информация в виде генетического кода хранилась в ДНК, белки состояли из; 20 аминокислот, энергия запасалась в виде АТФ и т. д.
Классификацию прокариот традиционно проводят по последовательностям гена 16S рРНК. Из проб, взятых в разных местах (например, из почвы, горячих источников или донных морских отложений) выделяют все имеющиеся там версии гена 16S рРНК и строят по ним эволюционные деревья. На деревьях часто обнаруживаются ветви, не соответствующие ни одной из известных групп прокариот.
|
|
|
|
Что интересно, клеточная мембрана у археобактерий и эубактерий возникла независимо.
[Может быть, эубактерии возникли на Земле в кислородной атмосфере? А археобактерии вообще могли прийти из космоса.]
Микоплазмы являются отдельным классом микроорганизмов, отличающимся как от вирусов, так и от бактерий. Они не имеют клеточной стенки [может быть, потеряли?] и ограничены плазматической мембраной. Неподвижны [как грибы]. Сапрофиты или паразиты. Это самые мелкие из существующих в природе организмов [за исключением нанобактерий?], способных самостоятельно жить и размножаться. Точно так же, как вирусы, микоплазмы не могут существовать иначе, чем паразитируя [противоречие - значит они не могут самостоятельно жить] на клетках хозяина. Микоплазмы способны расти на искусственных питательных средах, размножаются делением и почкованием.
В группу микоплазм входят два рода микроорганизмов - собственно микоплазма (Mycoplasma hominis, Mycoplasma genitalium) и уреаплазма (Ureaplasma urealiticum). Патогенные микоплазмы вызывают болезни человека (например, пневмонию, половые), животных (например, поражают легкие) и растений.
Риккетсии (Rickettsiaceae) — семейство бактерий. Названы по имени X. Т. Риккетса (1871—1910), в 1909 впервые описавшего возбудителя пятнистой лихорадки Скалистых гор. В том же году сходные наблюдения были сделаны Ш. Николем и его коллегами при исследовании сыпного тифа. В 1910 Риккетс погиб от сыпного тифа, изучением которого занимался в Мексике. В честь заслуг ученого возбудители этих инфекций были названы «риккетсиями» и выделены в род Rickettsia.
Типичный род Rickettsia представлен полиморфными, чаще кокковидными или палочковидными [как грибобактерии], неводвижными клетками. Грамотрицательны [?]. В оптимальных условиях клетки риккетсий имеют форму коротких палочек размером в среднем 0,2—0,6 ? 0,4—2,0 мкм, почти такие же, как и наиболее крупные вирусы (около 0,3 мкм). [Уже известны гигантские вирусы с 1 микрон. Сами риккетсии оказываются чуть крупнее нанобактерий.]. Их форма и размеры могут несколько меняться в зависимости от фазы роста (логарифмическая или стационарная фазы). При изменении условий роста они легко образуют клетки неправильной формы или нитевидные.
Нуклеоид клетки риккетсий содержит кольцевую хромосому. Размножаются путем бинарного деления, обладают независимым от клетки-хозяина метаболизмом. Источником энергии у внеклеточных риккетсий служит глутамат. Возможно, что при размножении получают макроэргические соединения из клетки-хозяина. Способны индуцировать [как?] свой фагоцитоз эукариотной клеткой.
На поверхности мембраны клеточной стенки располагается капсулоподобный слизистый покров и микрокапсула,
содержащие группоспецифичный «растворимый» антиген.
В клеточной стенке локализуются основные белки, большинство из которых являются видоспецифичными антигенами,
а также липополисахарид и пептидогликан.
В цитоплазматической мембране преобладают ненасыщенные жирные кислоты, она осмотически активна,
имеет специфическую транспортную систему АТФ-АДФ.
(Из Википедии)
Актиномицеты (Actinomicetes) или лучистые грибки, стрептомицеты, микобактерии (Mycobacterium - от греч. mykes - гриб и бактерии) - группа микроорганизмов, соединяющая в себе черты бактерий [эубактерий?] и грибов - всего 700 видов.
Распространены в почве, водоемах, в воздухе и на растительных остатках; некоторые - паразиты животных, человека (туберкулез, дифтерия и др.) и растений. Некоторые виды образуют антибиотики, пигменты, витамины [т.е. полезные]. Для них характерно нитевидное или палочковидное и кокковидное строение и наличие боковых выростов.
Актиномицеты состоят из центрального "клубка" ветвящихся нитевидных структур (гифы), от которого к периферии отходят тонкие филаменты. Длинный ветвящийся мицелий актиномицетов не имеет перегородок, чем сильно отличается от мицелия грибов.
Микобактерии, к которым относятся возбудители туберкулеза и проказы, обладают рядом особенностей, из-за которых с ними трудно бороться. Например, при лечении туберкулеза приходится принимать антибиотики очень долго, чтобы избежать рецидива, хотя большинство туберкулезных палочек (Mycobacterium tuberculosis) погибает в самом начале лечения. Дело в том, что некоторая часть популяции сохраняет жизнеспособность еще долго после гибели основной массы бактерий. Самое интересное, что выжившие микробы могут генетически ничем не отличаться от погибших. Иными словами, у микобактерий имеется большая ненаследственная изменчивость по устойчивости к антибиотикам. Микобактерии фактически создают фенотипическое разнообразие при каждом делении, не меняя своего генома.

Цианобактерии, или сине-зелёные водоросли (лат. Cyanobacteria, от греч. kyanos — сине-зелёный) — значительная группа крупных грамотрицательных эубактерий (около 2000 видов), способных к фотосинтезу, сопровождающемуся выделением кислорода. Сине-зеленая окраска обусловлена пигментами - хлорофиллом и фикоцианином. Размножение бесполое. Обитают чаще в пресных водах, но могут жить в морях, океанах, почве, горячих источниках. Некоторые съедобны. Цианобактерии, вместе с хлороксибактериями, относят к подцарству оксифотобактерий. Эти бактерии имеют одиночные и колониальные формы. Колонии создают органогенные известковые постройки (строматолиты). Цианобактерии могут использовать как солнечную энергию (автотрофность), так и энергию, выделяющуюся при расщеплении готовых органических веществ (гетеротрофность). В периферической части клеток цианобактерий диффузно распределены синий и бурый пигменты, определяющие в сочетании с хлорофиллом сине-зеленый цвет этих организмов. Некоторые цианобактерии могут иметь дополнительные пигменты, изменяющие их характерный цвет до черного, коричневого, красного. Цвет Красного моря определяется широким распространением в нем пурпурно пигментированных сине-зеленых. Цианобактерии наиболее близки к древнейшим микроорганизмам, остатки которых (строматолиты, возраст более 3,5 миллиардов лет) обнаружены на Земле. Они были и остаются самой распространенной группой организмов на планете. Сравнительно крупные размеры клеток и физиологическое сходство с водорослями было причиной их рассмотрения ранее в составе водорослей («синезелёные водоросли», «цианеи»). За то время было альгологически описано более 1000 видов в почти 175 родах. Бактериологическими методами в настоящее время подтверждено существование не более 400 штаммов. Биохимическое, молекулярно-генетическое и филогенетическое сходство цианобактерий с остальными бактериями в настоящее время подтверждено солидным корпусом доказательств, однако до сих пор некоторые ботаники, отдавая дань традиции, склонны относить цианобактерии к водорослям. |
Единственные, наряду с прохлорофитами, бактерии, способные к оксигенному фотосинтезу, предки цианобактерий рассматриваются в теории эндосимбиогенеза как наиболее вероятные предки хроматофоров красных водорослей (прохлорофиты по этой теории имеют общих предков с хлоропластами прочих водорослей и высших растений).
Сине-зелёные водоросли выделяют свободный кислород, одновременно химически связывая водород и углерод. Они замечательны тем, что способны использовать атмосферный азот и превращать его в органические формы азота. При фотосинтезе они могут использовать углекислый газ как единственный источник углерода. В отличие от фотосинтезирующих бактерий, цианобактерии при фотосинтезе выделяют молекулярный кислород. В течении прошедших 3-х миллиардов лет до начала кембрия они являлись основным источником свободного кислорода в атмосфере Земли, наряду с фотохимическими реакциями в верхних слоях атмосферы.

Строматолиты (др.-греч. στρῶμα «подстилка» и λίθος «камень», каменная подстилка, каменная прослойка) — ископаемые карбонатные (чаще известковые или доломитовые) стяжения, образовавшиеся на дне мелководного водоёма и представляющие собой ископаемые остатки цианобактериальных матов. Следует иметь ввиду, что вещество, из которого построен строматолит, не создается матом; последний лишь структурирует естественное осадконакопление. На ранних стадиях изучения строматолиты ассоциировались с остатками многоклеточных эукариот — губками, кораллами или мхами. По степени сложности они более всего напоминали исследователям скелеты многоклеточных эукариот. Позже к числу возможных строматолитообразователей были отнесены миксомицеты. Дальнейшее изучение строматолитов позволило однозначно связать их образование с жизнедеятельностью колоний нитчатых цианобактерий. Это было показано в результате обнаружения остатков нитей в ископаемых строматолитах и подтверждено исследованиями их современных аналогов. Строматолиты чаще всего состоят из карбоната кальция потому лишь, что карбонатный тип осадконакопления в море наиболее обычен, однако в иных гидрохимических условиях формируются строматолиты фосфатные, кремнеземовые, железистые и пр. Мат, располагающийся на верхней поверхности создаваемого строматолита, представляет собой плотный многослойный "ковер" общей толщиной до 2 см; основу его составляют нитчатые либо пальмеллоидные цианобактерии, однако помимо них в формировании сообщества участвуют и другие бактерии. Маты существуют во многих районах мира, однако в современное время настоящие строматолиты существуют только в Акульем заливе на западном побережье Австралии и на атлантическом побережье Багамских островов. Многослойная расцветка строматолитов может меняться в течении суток, поскольку обитатели нижних слоев могут подниматься в темное время наверх и наоборот. Скользят бактерии вверх и вниз со скоростью до 2см в час. Строматолиты достоверно появляются в геологической летописи в древнейших осадочных формациях Уарравуна (Западная Австралия) возрастом в 3,5 млрд лет – это древнейшая известная форма [прокариотической] жизни. Наибольший расцвет цианобактерий пришелся на протерозойский эон, затем их роль резко снизилась. Строматолиты обитали в соленых и пресных водах. В протерозое из строматолитов состояли огромные рифы мощностью в сотни метров. Отдельные глубоководные строматолиты достигали высоты 75 м. Протерозойские строматолиты достигли высокого уровня сложности: появились формы со всевозможными ветвящимися столбиками, козырьками, разнообразной слоистостью и микроструктурой и т. п. Современные строматолиты, образуемые бактериальными матами, устроены намного проще. |

Министроматолиты - мельчайшие столбчатые строматолиты с диаметром столбиков <20 мм - появились в геологической летописи 2.9 млрд. лет назад и стали довольно многочисленными 2.8-2.5 млрд. лет назад. Систематический состав их архейских представителей изучен крайне фрагментарно: валидно описаны 4 формальных рода и 6 формальных видов, из которых 2 рода и 1 вид поднимаются в нижний протерозой. Представительный комплекс раннепротерозойских министроматолитов имеет возраст 2.3-1.65 млрд. лет и включает 41 формальный вид и 16 формальных родов, которые по наиболее крупным морфологическим особенностям разделяются на 5 надродовых группировок. Следующий возрастной комплекс министроматолитов, развитый в раннем и начале среднего рифея (1.65-1.20 млрд. лет назад), включает 62 формальных вида и 15 формальных родов, из которых только 3 рода и 1 вид объединяют его с раннепротерозойским. В целом рифейские министроматолиты однообразнее раннепротерозойских из-за исчезновения одной сложно построенной надродовой дорифейской группировки, преобладания в рифее форм с цилиндрическими вертикальными колонками и появления короткостолбчатых построек, связанных протяженными наслоениями. Наряду с этим, рифейские министроматолиты проявляют явную тенденцию к уменьшению диаметра и высоты колонок и к увеличению количества переходных мостиков. [Вендские строматолиты возрастом около 620 млн. лет называют онтолитами. Имеются и возрастом 775 млн. лет.] Во второй половине среднего рифея министроматолиты практически исчезают из геологической летописи [хотя известны остатки и позднепротерозойских строматолитов возрастом около 1 млрд лет], но вновь появляются в ней в раннем палеозое. Тенденции морфологических изменений министроматолитов, зафиксированные в протерозое, не находят продолжения в их раннепалеозойском комплексе и не совпадают с тенденциями изменения протерозойского комплекса столбчатых строматолитов обычной размерности. Поэтому можно предполагать, что ответственность за формирование каждого из упомянутых комплексов несли специфические ассоциации микроорганизмов. |
Прохлорофи́ты (лат. Prochlorales — «дохлорофильные дробянки») — порядок прокариот, обычно относимый к царству бактерий, отличительной особенностью представителей которого является способность к оксигенному фотосинтезу, сходному с таковым у цианобактерий при отличном от цианобактерий составе фотосинтезирующих пигментов. Возможно, вместе с цианобактериями участвовали в строительстве строматолитов.
В силу своей редкости прохлорофиты не имеют какого-либо существенного практического значения, однако представляют немалый научный интерес как возможные «предки» хлоропластов эукариот. Предполагается, что симбиоз каких-то других прокариот с прохлорофитами дал начало зеленым водорослям - предкам многоклеточных растений.
От гипотетических протобионтов следует строго отличать археобактерии (археи). Недавно они были признаны отдельной самостоятельной группой. Они настолько отличаются от всех остальных живых существ, что представляют собой целый "мир", отдельный от других бактерий (эубактерий) и организмов с ядросодержащими клетками (эукариотов). Они отличаются своим способом выживания в экстремальных условиях: выдерживают под давлением нарастающую температуру до 110°С и рН-величины от 1. Кроме того, это некультивируемые микробы, отказывающиеся расти на лабораторных средах.
Царство архей (ранее архебактерии), впервые описано в 1977 г.: липиды состоят из соединений глицерина и терпенов, мембраны монослойные липидные, клеточная стенка построена из псевдомуреина или из гликопротеидов, или из кислых полисахаридов, ДНК-зависимая РНК-полимераза состоит из 9-11 субъединиц, в тРНК нет риботимидина.
Археи — чрезвычайно разнообразная группа, однако значительная часть их разнообразия известна лишь по последовательностям гена 16S рРНК, по которому строится эволюционное дерево прокариот. Ветвь архей - это новая, неизвестная группа, степень родства которой с известными микробами можно определить только приблизительно, так как одного-единственного гена 16S рРНК недостаточно для более строгих выводов.
С точки зрения эволюционной теории эти существа подходят в качестве кандидатов, которые в наибольшей степени похожи на предполагаемые первые живые формы, т.к. окружающая их среда, вероятно, наиболее близка условиям раннего периода развития "остывающей" Земли с многочисленными вулканическими процессами. В эту картину хорошо вписывается и встречающееся у некоторых особей, рассматриваемое как "первичное", серное дыхание (энергия выделяется при распаде H2S на Н2, и S). Поэтому эту группу и назвали "археобактерии" (древние бактерии).
Есть мнение, что от археобактерий произошли независимо как эубактерии, так и эукариоты [серобактерии, азото-фосфорные бактерии, "экстремальные" микробактерии и ультрамикробактерии - археи?].
Царство архебактерий представляет собой своеобразную и наименее изученную таксономическую группу прокариот. По своей морфологии Archeabacteria похожи на эубактерии, но на молекулярном уровне они сближены с эукариотами. Эти микроорганизмы рассматривают как прокариотические эволюционные предшественники эукариот.
Архебактерия Methanococcus jannaschii, первичная структура генома которой была определена в 1996 г., обнаружена в горячих морских глубоководных источниках. Энергию для жизнедеятельности этот микроорганизм получает при восстановлении двуокиси углерода до метана молекулярным водородом. Температура, близкая к температуре кипящей воды, является оптимальной для его роста, который может происходить при давлении более 200 атм. M. jannaschii не требует для своего роста органических соединений: все необходимое для жизни он синтезирует из неорганических веществ.
Геном M. jannaschii состоит из основной кольцевой хромосомы и двух небольших внехромосомных элементов, размеры которых составляют соответственно 1700, 58 и 16 т.п.о. Подобные размеры геномов типичны для архе- и эубактерий. GC-состав ДНК этого термофила невысок и составляет всего 31%. Геном организован компактно: обнаружено около 1700 потенциальных кодирующих участков ДНК, по одному на каждые 1000 п.о. Многие ДНК-локусы M. jannaschii не обнаруживают гомологии с известными последовательностями. Функциональное значение большого числа потенциальных кодирующих последовательностей генома этого микроорганизма остается невыясненным.
M. jannaschii отличается от других прокариот и эукариот большим набором только ему свойственных генов и функций. Анализ структуры генома M. jannaschii показал, что гены, организующие системы обработки генетической информации - транскрипции, трансляции и репликации ДНК, в большей степени напоминают гены эукариот, чем бактерий... Таким образом, система транскрипции архебактерий представляется как более простая версия соответствующей эукариотической системы...
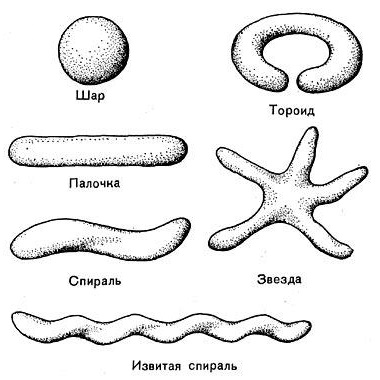
Эубактерии (лат. Eubacterium) — род грамположительных [?] строго анаэробных бактерий. В строгом смысле к истинным бактериям относят одноклеточных бактерий с ригидными клеточными стенками, гетеротрофов, перемещающихся (если обладают подвижностью) с помощью жгутиков. В эту группу входит преобладающее количество прокариотных микроорганизмов. [Большинство] не образуют спор. Форма чаще всего палочковидная. Отличаются жёсткой клеточной стенкой. Эубактерии (Царство Eubacteria, собственно бактерии, греч. eu — хорошо и bacterion — палочка) - истинные бактерии, класс бактерий, в который входят организмы с неветвящимися клетками и прочными клеточными стенками, имеющие форму мелких тонких палочек, реже кокков, среди них находятся также нитевидные и спиралевидные формы. Часть эубактерий обладает подвижностью, часть - неподвижны; движение осуществляется при помощи жгутиков простого строения, состоящих из взаимно перевитых фибрилл. Часть эубактерий способна формировать эндогенно (внутри клеток) термоустойчивые споры, большинство же представлено неспороносными грамотрицательными [?] формами. Размножаются эубактерии делением, некоторые нитчатые бактерии образуют специальные репродуктивные клетки - гонидии. Клетки эубактерий очень малы и вместе с тем сильно различаются по размерам: от карликовых форм (диаметром около 0,125 мкм) до гигантов, длиной до 10 мкм (Ghromatium okenii). В природных субстратах (илах, почвах) можно встретить стебельковых бактерий, достигающих в длину вместе со стебельком 100 мкм и более. Следовательно, гиганты превышают карликов по размерам в 100 и 1000 раз. Большинство палочковидных бактерий по длине не превышают 5 мкм, по толщине 1 мкм. У многих псевдомонад диаметр клетки равен 0,4—0,7 мкм, у ряда почвенных форм — 0,2—0,3 мкм. Существующие сейчас классификации эубактерий довольно сложны и основаны на разных критериях: типу питания и обмена веществ, молекулярным особенностям, экологии и т.д. А, если по-простому, класс Eubacteriae объединяет порядки: Eubacteriales (эубактерии), Chlamydobacteriales (хламидобактерии), Ferribacteriales (железобактерии), Thiobacteriales (серобактерии). |
Ключевые слова для поиска сведений о доядерных формах жизни (организмах-прокариотах):
На русском языке: прокариоты, доядерные формы жизни, безъядерные микроорганизмы,
простейшие организмы без ядра, микробы, пробионты, протисты, бактерии,
сине-зеленые водоросли, цианобактерии, археобактерии, эубактерии, археи, рикетсии,
микроскопические грибы, грибобактерии, актиномицеты, стрептомицеты, микобактерии, микоплазмы;
На английском языке: prokariota, procaryota.
|
|
|
|
|
|