|
|
Главная |
|
|
Пишите |
|
|
"...Бог следит внимательно за теми процессами, которые происходят на Земле, и пока эти процессы развиваются в соответствии с задуманным сценарием он остаётся сторонним наблюдателем. Но стоит эксперименту принять нежелательный поворот, как следует оперативное вмешательство." (Лимарев) |
Эукариоты ("настоящие ядерные") - организмы, с обособленным в ядре генетическим аппаратом [какие это даёт преимущества?]. Кроме того, они имеют различнвые органеллы с собственным генетическим материалом.
Эукариотами являются как одноклеточные протисты, так и все многоклеточные организмы (Metazoa): растения, грибы и животные [хотя среди протист тоже есть организмы со всеми этими типами обмена веществ и их комбинациями]. Все эти группы организмов имеют общий план строения клеток и, по современным представлениям, общее происхождение. Поэтому группа ядерных рассматривается как монофилетический таксон наивысшего ранга.
Согласно наиболее распространённым гипотезам, эукариоты появились 1,5–2 млрд. лет назад (в среднем или раннем протерозое). [Вероятно, во время одного из спадов строматолитов во время гляциоэры.] Важную роль в эволюции эукариот сыграл симбиогенез — симбиоз между эукариотической клеткой, видимо, уже имевшей ядро и способной к фагоцитозу (поеданию), и проглоченными этой клеткой бактериями — предшественниками митохондрий и пластидов.
По-видимому, была проглочена и "осимбионена" и цианобактерия - так у протопротист появились хлоропласты, и они изначально были (вернее, стали) "гриборастениеживотными":
К слову сказать, возможно, что симбионты поначалу просто паразитировали в теле археобактерий.
Т.к. объекты симбиоза могли быть разными, то, возможно, почти все протисты - полифилетические, т.е. имеют разных предков-симбионтов.
Разделы страницы о появлении и особенностях эукариотических одноклеточных организмов:
Также смотрите страницу о путях превращения одноклеточных организмов в многоклеточных животных (Metazoa):
Эукариоты обладают 2-мя универсальными белками — актином и миозином, обеспечивающими разнообразные типы клеточной подвижности: амёбоидную активность, движение органелл внутри клетки, а у высших организмов — мышечные сокращения. Актиново-миозиновая система позволяет образовывать псевдоподии, захватывать ими жертву и формировать пищеварительные вакуоли (даже вирусы проникают в клетку эукариот путём провокации так называемого „эндоцитоза“ — клетка принимает их за нечто полезное, „проглатывает“, и вирус, оказавшись в цитоплазме, начинает свою разрушительную работу). Приобретение актиново-миозиновой системы позволило эукариотам питаться путём фагоцитоза, активно захватывая крупные пищевые частицы. [Интересно, а как же питаются бактерии-гетеротрофы - разве они не "захватывают" пищу?]
Короче, принципиальный мотив происхождения и источник отличий эукариот от прокариот - хищный образ жизни. Одноклеточные эукариоты - это гениальные акулы среди микроорганизмов. Сначала у них появились "мускулы" (актиново-миозиновая система). Они стали активно двигаться и поедать бедных бактерий. Какие-то из них были пощажены и использованы как инструмент. Особо полезными оказались цианобактерии со своим фотосинтезирующим аппаратом. Фотосинтезирующие пластиды стали аккумулятором энергии, который давал возможность двигаться в поисках пищи до тех пор, пока она не будет найдена. Вопрос - не появилась ли в связи с таким активным образом жизни и особая система чувствительности? А раз так, то и растения тоже чувствуют?
Перечислим основные отличия между эукариотическими (ядерными) и прокариотическими (доядерными) организмами:
Кроме того, у бактерий обмен веществ разнообразнее, т.к. среди них встречаются все 4 типа питания (фотоавтотрофное, фотогетеротрофное, хемоавтотрофное, хемогетеротрофное). Эукариоты же или сами синтезируют энергию из солнечного света, или берут готовую энергию такого происхождения. Это может быть связано с появлением у эукариот хищников, необходимость синтезировать энергию у которых отпала.
Как следствие большей структурной сложности, эти 2 группы организмов сильно отличаются по своим средним размерам. Диаметр прокариотической клетки составляет обычно 0,5–10 мкм, а у эукариот - 10–100 мкм. Т.о. объём эукариотической клетки в 1000–10000 раз больше, чем прокариотической. Рибосомы прокариот мелкие (70S-типа). А клетки эукариот содержат как более крупные рибосомы 80S-типа в цитоплазме, так и 70s-рибосомы прокариотного типа в митохондриях и пластидах.
Эукариотные организмы — простейшие [протисты], различные группы растений, грибы и многоклеточные животные — доминируют в современной биосфере. Однако все они несут в своих клетках симбионтов — потомков древних свободноживущих бактерий. Только благодаря им эукариотные организмы способны жить в кислородной атмосфере и использовать энергию солнечного света для синтеза органических веществ. Так, может быть, на самом деле, эукариоты вовсе не доминируют в биосфере?
На сегодняшний день твердо установлено, что митохондрии и пластиды эукариотической клетки являются потомками симбиотических бактерий - альфапротеобактерий (кислорододышащих) и цианобактерий, соответственно. Этот симбиоз возник более двух миллиардов лет назад. Митохондрии были уже у последнего общего предка всех современных эукариот: это их универсальная черта. Хотя некоторые современные эукариоты лишены митохондрий, это — результат вторичной утраты.
В 1967 году, Линн Маргулис (Lynn Margulis) опубликовала развернутое изложение симбиогенетической теории, согласно которой эукариоты (организмы с клеточными ядрами) возникли в результате серии объединений разных клеток между собой.
Современная поправка к этой теории гласит, что в основе становления эукариот, по-видимому, была не общая тенденция, охватившая многие эволюционные ветви (как предполагала Маргулис), а уникальное событие, приведшее к слиянию клеток археи и протеобактерии. В результате образовалась сложная клетка с митохондриями, которая и стала первым эукариотом.
Дальнейшие симбиогенетические события — например, захват водорослей, ставших хлоропластами, — действительно происходили много раз, но с возникновением эукариот как таковых они не связаны.
Американский биолог Л. Томас сказал: „Обычно на митохондрии смотрят как на порабощённые существа, взятые в плен, чтобы снабжать АТФ клетки, и не способные дышать самостоятельно. Но с точки зрения самих митохондрий они — существа, которые давным-давно нашли для себя лучшее из возможных пристанищ, где можно жить, затрачивая минимум усилий и подвергаясь наименьшему риску“. Поэтому мы не должны забывать, что в каждой клеточке нашего тела живут крошечные потомки древних оксифильных [кислородолюбивых] бактерий, которые прокрались в организм наших далёких предков 2 млрд. лет назад и продолжают существовать в нас, сохраняя собственные гены и свою особую биохимию.
Какие же прокариоты могли быть предками митохондрий? Среди современных прокариот ближе всего к ним пурпурные альфа-протеобактерии (об этом говорят, в частности, и новые данные молекулярной филогении) — аэробные фотосинтезирующие бактерии, мембрана которых образует глубокие впячивания, похожие на кристы митохондрий. Их прародители, вероятно, обитали в кислородных карманах анаэробной биосферы. Вступив в симбиоз с древними амёбоидными эукариотами, протеобактерии утратили способность к фотосинтезу, т.к. все необходимые органические вещества стали получать от хозяина — древнего эукариота, получившего тоже свою выгоду: он перестал бояться высоких концентраций кислорода, который утилизировали симбионты. [Видимо, толчком к такому симбиозу послужила "кислородная катастрофа" в начале протерозоя].
По наиболее популярной (трёх-симбионтной) версии, предком эукариот было сообщество прокариот, включавшее как минимум 3 компонента:
А согласно альтернативной (2-компонентной) версии, предки эукариот были бисимбионтами из археи (метаногенной) и простейшего грибообразного (спорообразующей клостридии).
Что касается 3-го компонента, то приобретение фотосинтезирующих симбионтов стало одним из магистральных направлений эволюции первичных эукариот. Предполагается, что они вначале были одноклеточными животными и питались, захватывая и переваривая другие микроскопические организмы. А включенные симбионты превратилились в органеллы, обеспечившие синтез органических веществ из углекислого газа и воды за счёт энергии солнечного света. Этот путь привёл к появлению различных групп эукариотных растений, то есть автотрофных фотосинтезирующих организмов. Они не родственны друг другу и возникли в результате симбиоза хищных протистов (простейших или их колоний) с различными фотосинтезирующими организмами.
Симбионтами хищных эукариот в нескольких случаях стали цианобактерии — сине-зелёные водоросли, самая распространённая (по крайней мере в современной биосфере) и, возможно, самая древняя группа фотосинтезирующих прокариот. Их несомненными потомками являются фотосинтезирующие органеллы (хлоропласты) багрянок. Они окружены только двумя мембранами, имеют собственную кольцевую ДНК и рибосомы прокариотного типа и содержат типичные для цианобактерий хлорофилл „а“ и специфические пигменты цианобактерий — фикобилины.
Багрянки (красные водоросли) в настоящее время широко распространены в морях нашей планеты. Они способны существовать на глубинах в несколько сот метров, но живут и в приливно-отливной полосе, а некоторые виды обитают и в пресных водах. Возможно, красные водоросли — самая древняя группа эукариотных растений. Об этом говорит полное отсутствие в их жизненном цикле жгутиковых стадий (даже их сперматозоиды — безжгутиковые), что позволяет предположить, что предки этих водорослей отделились от остальных эукариот ещё до приобретения жгутиков.
|
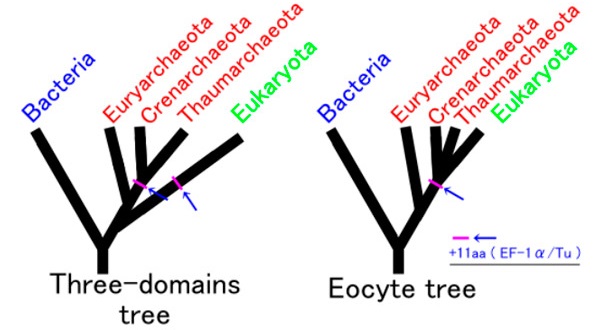
Трехдоменный (слева) и двухдоменный варианты древа жизни. Согласно классической трехдоменной версии, эукариоты отделились от общих с археями предков до того, как началось расхождение групп современных архей (эвриархей, кренархей и др.). Согласно двухдоменной версии, эукариоты — одна из веточек архей, обособившаяся уже после начала их дивергенции. |
В донных осадках на глубине 3283 м в Северном Ледовитом океане обнаружены микроорганизмы из надцарства архей,
более близкие к эукариотам, чем любые другие прокариоты.
Судя по набору генов, группа новооткрытых в 2015 году микробов, названная локиархеями,
обладает многими признаками эукариот, включая актиновый цитоскелет и способность к фагоцитозу.
Скорее всего, именно благодаря этой способности предкам эукариот удалось захватить ["съесть"] бактериального симбионта, давшего начало митохондриям.
Это говорит о том, что эукариоты - не родственники археобактерий, а одна из их групп.
Природа «хозяйской» клетки, некогда захватившей бактериальных симбионтов, менее очевидна, чем происхождение митохондрий и пластид. Геном эукариот явно имеет химерное происхождение: часть генов досталась от архей, другая — от бактерий (в т.ч. от симбионтов). Гены архейного происхождения выполняют в основном «центральные» функции (работа с генетической информацией и синтез белка), гены бактериального происхождения — в основном «периферические» (обмен веществ, взаимодействие с внешней средой). По-видимому, предок эукариот (та клетка, которая приобрела митохондриального симбионта) был близок к археям, а бактериальных генов он нахватался путем горизонтального переноса. Недавние исследования показали, что массированное заимствование бактериальных генов происходило на ранних этапах формирования чуть ли не всех крупных клад архей, так что предок эукариот не был исключением. Но эукариотическая клетка устроена намного сложнее, чем прокариотическая, причем далеко не для всех эукариотических генов и молекулярных систем найдены очевидные прокариотические предшественники или аналоги. Откуда взялись эти уникальные особенности эукариотической клетки — вот главный вопрос, на который необходимо ответить.
Одной из особенностей геномов локиархей является повышенная доля генов бактериального происхождения. Это согласуется со сказанным о периодическом массовом заимствовании бактериальных генов археями, в т.ч. и предками эукариот. Если локиархеи — ближайшая родня эукариот, то у них могут найтись гены и признаки, характерные для эукариот, но отсутствующие у других прокариот. И действительно, в геноме Lokiarchaeum нашлось целых 175 белок-кодирующих генов (3,3% от общего числа генов в геноме), похожих на эукариотические, в т.ч. на т.н. ESPs (eukaryotic signature proteins) — белки, считающиеся уникальными для эукариот. Это, наряду с эволюционным деревом, — второй весомый довод в пользу близкого родства локиархей и эукариот.
Целенаправленный поиск микробов, родственных локиархеям, привел к обнаружению большой и разнообразной группы, заслуживающей ранга надтипа и получившей название «Асгард». В эту группу, помимо описанных ранее локиархей и торархей, входят два новых типа: одинархеи и хеймдалльархеи.
В геномах асгардархей закодировано множество белков, ранее считавшихся характерными только для эукариот, в том числе многие компоненты цитоскелета, а также молекулярные системы, отвечающие за везикулярный транспорт и убиквитин-зависимую деградацию белков.
Реально (в виде организмов) асгардархей еще не нашли, но геномные данные указывают на более сложную организацию их клеток по сравнению с обычными прокариотами. Предки эукариот почти наверняка были асгардархеями, возможно, близкими к хеймдалльархеям. Открытие показало, что ряд шагов в сторону усложнения организации был сделан предками эукариот задолго до приобретения митохондриального симбионта.
Поскольку систематика эукариотических микроорганизмов лежит в основе систематики многоклеточных, будем в этом разделе рассматривать теории и экспериментальные данные по генетической группировке живого мира.
Профессор эволюционной биологии Оксфордского университета Thomas Cavalier Smith предлагает следующую классификацию протистов:
Простейшие
|
-------------------
| |
Эубактерии Neomura
|
-------------------------------
| |
Археи Эукариоты
|
------------------------------------------------------
| |
Uniconta Biconta
| |
-------------- ---------------------------------
| | | | |
Амёбы Opisthokonta Cabozoa Apusozoa |
| | |
----------------- ------------------- -------------------
| | | | | |
Грибы | Excavata Rhizaria Растения Chromalveolata
---------------- | |
| | ----------------- -----------------
Животные Choanozoa | | | |
Cercozoa Retaria Хромисты Alveolata
По новейшим данным шведско-норвежских генетиков, эукариоты делятся не на 5, а на 4 царства (похоже на оксфордскую классификацию):
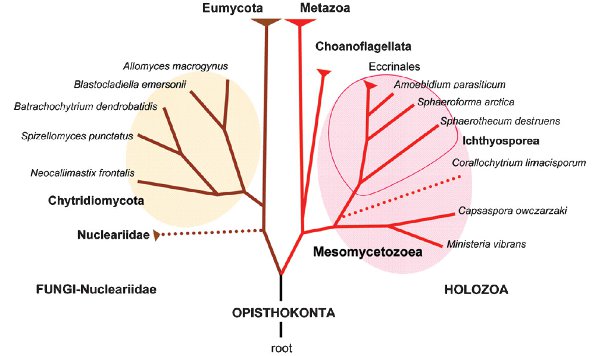
На рисунке ниже показана классификация опистоконт в виде эволюционного дерева (2009 г.). Opisthokonta (заднежгутиковые) — общее название эволюционной ветви, к которой относятся животные, воротничковые жгутиконосцы, мезомицетозои и грибы, которые иногда объединяют в парафилетический тип Choanozoa.
Одной общей чертой всех Opisthokonta является то, что жгутиконосные клетки, такие как сперматозоиды большинства животных и споры хитридиомицет, передвигаются при помощи одного заднего жгутика. Это дало группе своё название. В отличие от них, другие эукариоты перемещаются при помощи одного или нескольких передних жгутиков.
Считается, что многоклеточные животные произошли из колоний воротничковых жгутиконосцев (хоанофлагеллятов), но к ним близки и многоядерные мезомицетозои.
|
Непосредственный предок многоклеточных животных пока не известен. По каким-то признакам, он мог быть ближе к мезомицетозоям (Mesomycetozoea), чем к воротничковым жгутиконосцам (Choanoflagellata). Последние хоть и близки к животным (Metazoa), но далеко не во всем на них похожи, они явно успели пройти собственную длинную эволюцию. Нет сомнений в том, что группа (Metazoa + Choanoflagellata + Mesomycetozoea) в целом является тесной и единой. Она характеризуется, например, несколькими общими уникальными генами. В современных работах эту группу называют Holozoa («все животные»).
Из 5000 исследованных генов ученые выделили 123 общих для всех известных групп организмов, которые были затем подробно изучены. Все небактериальные (высшие эукариотические) формы жизни на Земле можно теперь разделить на 4 основные группы вместо 5.
Науке, скорее всего, до сих пор не известны все дожившие до наших дней супергруппы эукариот. Найти новую «рано ответвившуюся» линию, радикально отличающуюся от всех известных, можно даже в таких доступных и, казалось бы, хорошо изученных системах, как микробные сообщества, ассоциированные с тропическими кораллами. Поиск и изучение подобных организмов сможет пролить новый свет на такие запутанные вопросы, как ранняя эволюция эукариот в целом и их митохондрий в частности. Например, изучение анкорацисты показало, что нельзя судить о порядке ветвления древнейшей (базальной) части эволюционного дерева эукариот по таким «многообещающим», как считалось, признакам, как наличие той или иной системы созревания цитохрома C и размер митохондриального генома.
Ключевые слова для поиска сведений об эукариотах - их эволюции и дивергенции:
На русском языке: эукариоты, ядерноклеточные микробы, настоящие клетки, организмы с ядросодержащими клетками, ядерные клеточные существа;
На английском языке: Eukariota, Eukaryota, Eucariota, Eucaryota.
|
|
|
|
|
|