|
|
Главная |
|
|
Пишите |
|
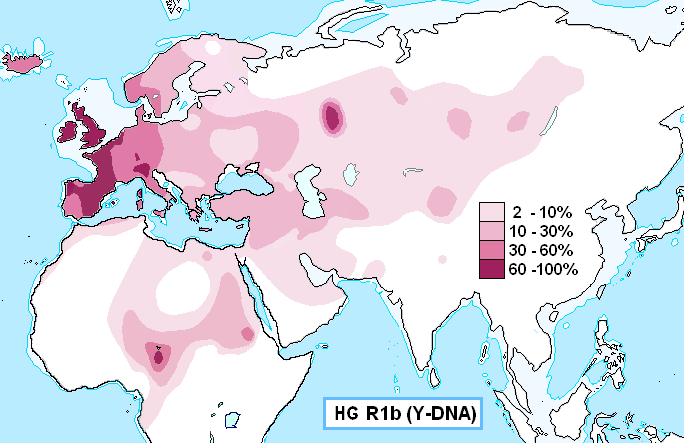
Гаплогруппа R1b является самой распространенной гаплогруппой в Западной Европе (особенно на Атлантическом и Северном побережье), достигая более 80% среди населения в Ирландии, Шотландии, Уэльс-западный, в Атлантической Франции и в Стране Басков [мореходы и строители мегалитов]. Также распространена в Анатолии и по всему Кавказу, в регионах России и в странах Центральной и Южной Азии. Максимумы - в долинах рек на севере центральной Италии (более 70%) [Тоскана? - страна этрусков], у осетин-дигорцев на Северном Кавказе (около 23%), в Армении (27%), у башкир на Урале (43%), в Туркменистане (30%), у хазарейцев Афганистана (25%), уйгур Северо-Западного Китая (до 20%). Субклад R1b-V88 широко распространен даже в экваториальной Африке, встречается до 95% у мужчин в северном Камеруне. Гаплогруппа R1b — гаплогруппа, наиболее распространённая в Западной Европе (субклад R1b1a2) и на Южном Урале. Встречается также в Центральной Азии, Восточной Европе, Северной Африке, в Арабских странах. После миграций европейцев в Америку и Австралию она составляет значительную долю и там. Типичные представители: потомки кельтов, баски, германцы, башкиры, камерунцы [!]. Предполагаемая дата появления гаплогруппы R1b: 16 500 лет до н. э. Предполагаемое место появления гаплогруппы R1b: Центральная Азия или Ближний Восток. Сестринские гаплогругруппы: R1a. Характеристика мутаций: M343. Определяется однонуклеотидным полиморфизмом M343, открытым в 2004 году. С 2001 по 2005 год R1b определяли наличием ОНП P25. В других системах классификации — Hg1 и Eu18. Гаплогруппа R1b является наиболее частотной гаплогруппой в Западной Европе (57%). В германоязычных странах наиболее частотна она в странах Бенилюкса (в Голландии 70.4%, у фризов 56%, в Бельгии 63%). В Германии средняя частотность этой гаплогруппы 47.9%, причем ее процент в южной Германии выше, чем в северной. В меньшей степени эта гаплогруппа характерна для Скандинавии, ср. норвежцы 25% (по другим данным 31.3%), шведы 24%, датчане 42.9% (Karlsson et al. 2006; Dupuy et al. 2006), причем на севере Скандинавии процент этой гаплогруппы ниже чем в центральной Скандинавии (Lappalainen et al. 2009). В Норвегии процент R1b уменьшается к востоку, ср. 45% носителей R1b на юго-западе Норвегии и 26% на востоке (Dupuy et al. 2006). У южных и особенно у западных соседей германоязычного населения Европы процент гаплогруппы R1b очень высок (у валлийцев - более 90%, у ирландцев и басков - более 80%, у французов и северных итальянцев - более 60%). Частотность этой гаплогруппы убывает по направлению на восток (у поляков 10–14%, по другим данным 22.2%, у латышей 15%, у литовцев 5%, у саамов около 6%, у финнов 2%). Что касается датировки появления R1b в Европе, то здесь мы сталкиваемся с двумя гипотезами. Если у генетиков фактически нет разногласий относительно ее азиатского происхождения и времени ее появления (около 20 000 до н.э. или чуть позже), то предположения о времени ее появления в Европе сильно отличаются друг от друга. Еще совсем недавно считалось, что все варианты R1b в Европе восходят к эпохе верхнего палеолита, т. е. к эпохе охотников и собирателей (Karlsson et al. 2006). В последнее время, однако, появление этой гаплогруппы в Европе стали относить даже не к мезолиту, а к неолиту, связывая движение R1b в Европе с миграцией населения из Западной Азии, которое принесло земледелие в Европу (Myres et al. 2010). Если мы посмотрим на распространение гаплогруппы R1b в Европе, то очевидно, что в исторический период она распространялась с запада на восток. Наиболее частотной она остается у самых западных народов (современные кельтоязычные и романоязычные народы западной Европы, а также баски). Распространение этой гаплогруппы у германоязычных народов с наибольшим ее процентом на западе германоязычной области несомненно свидетельствует о генетическом участии носителей гаплогруппы R1b (к которым прежде всего относятся предки современных кельтоязычных и романоязычных народов) в формировании генетического пула германоязычных народов. (Кузьменко Ю.К.) Предполагаемое направление миграций "эрбинов": Средняя Азия (верхний палеолит?) -> Южный Урал (мезолит?) -> Причерноморье (неолит? - строители дольменов в Адыгее) -> Анатолия (хатты - родственники адыгов - энеолит) -> Средиземноморье (морским путём), Пиренеи и (по побережью) Западная Европа (ранняя бронза - палеоевропейцы-мегалитники) -> Засахарье (конец бронзы - минойцы-гараманты после Троянской войны). Возможно, существовала и вторая, более ранняя, ветвь проникновения "эрбинов" в Европу: Средняя Азия (Туркмения) -> Ближний Восток (Сирия) -> Северное Средиземноморье (неолитическая культура кардиальной керамики) -> западно-атлантическое побережье Европы. Кстати, гараманты-критяне, совершившие переход Сахары на колесницах, имели собственную, по-видимому, слоговую эгейскую письменность, которая, возможно, повлияла на создание некоторых самобытных письменностей Африки. |
Разделы о клане эрбинов (К1b):
Имеются предположения о связи носителей группы R1b со строителями мегалитов, культурой колоколовидных кубков и палеоевропейцами, которые изучены и интегрированы автором в его исследовании. Возможно, причиной миграций этих популяций служили периодические колебания увлажненности степей Приуралья и Южной Сибири, совпадающие также с колебаниями уровня Каспийского моря.

Происхождение гаплогруппы R1b не совсем ясно и по сей день [см. гипотезу автора]. Некоторые из самых старых форм R1b находятся на Ближнем Востоке и в Закавказье. Гаплогруппа R1* и R2*, возможно, возникла на юге Центральной Азии (между Каспийским морем и Гиндукушем), либо в Южной Азии. Субклад R1 развился в R1b*, потом в R1b1* в северной части Ближнего Востока во время ледникового периода. Это произошло предположительно при переходе в северную Анатолию и на Кавказ в начале неолита, где появился субклад R1b1a. Ближневосточные остатки превратились в R1b1a (M18), который теперь встречается в низких частотах у ливанцев и у друзов. Финикийцы (которые пришли из современного Ливана) распространили эту гаплогруппу R1b1a и R1b1* в своих колониях, в частности, на Сардинии и в Магрибе. Британские генетики Брайан Сайкс и Стивен Оппенгеймер утверждали, что гаплогруппа R1b не имеет отношения к индоевропейскому заселению Западной Европы и происходит от палеолитического (доиндоевропейского) населения Иберии. Взгляды Сайкса и Оппенгеймера одно время имели широкое распространение в Европе благодаря написанными им популярным бестселлерам о генетической истории Европы. С другой стороны, подобный взгляд на происхождение R1b сталкивается с непреодолимыми противоречиями [?]. |
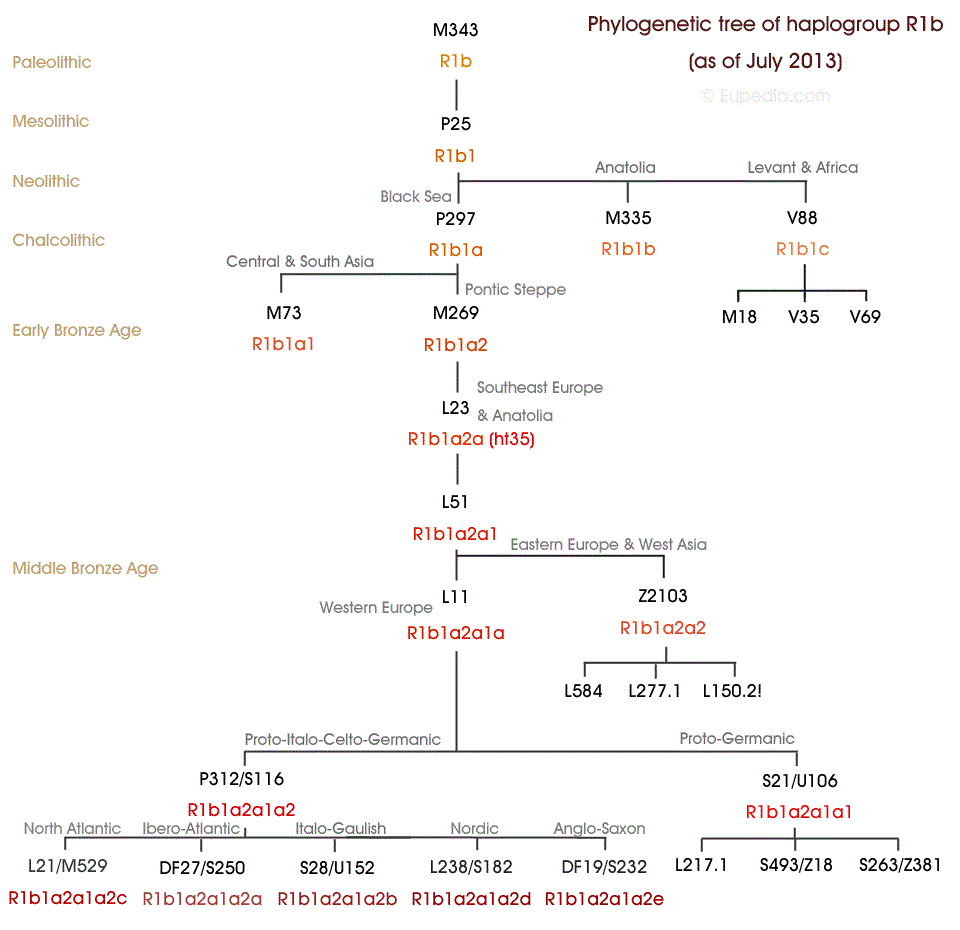
Последующие исследования установили, что разнообразие субкладов данной гаплогруппы увеличивается по мере движения на восток, что определенно говорит о восточном происхождении данной гаплогруппы R1b. Ряд современных генетиков полагают, что R1b зародилась в Центральной или Западной Азии. Сначала была выдвинута гипотеза, что R1b является коренной для Западной Европы, поскольку именно там она преобладает. Впоследствии было доказано, что гаплотипы R1b демонстрируют большее разнообразие малых побочных ответвлений в Анатолии и на Кавказе, чем в Европе. Также европейские субклады более молоды по сравнению со средневосточными или центральноазиатскими. Основная европейская ветвь R-P312/S116 или гаплогруппа R1b1a2 восходит всего лишь к 3500 или 3000 до Р.Х. Это не значит, что старейший общий предок этой линии прибыл в Западную Европу в этот период, но значит, что первый человек, нёсший мутацию R-P312/S116, жил по крайней мере 5000 лет назад, вполне вероятно, где-то в долине нижнего Дуная или в окрестностях Чёрного моря. В любом случае, эти временные рамки слишком малы для палеолитического происхождения или неолитического пришествия R1b. Открытие того, что считалось «европейскими линиями», в Средней Азии, Пакистане и Индии вбило последний гвоздь в идею палеолитического происхождения R1b в Западной Европе и подтвердило её связь с индоевропейцами Причем, история гаплогрупп R1b и R1a на начальном этапе были неразрывно связаны друг с другом. Субклады R1b1a1 и R1b1a2 (наиболее распространенные в Европе), тесно связаны с распространением индоевропейских языков, о чем свидетельствует их присутствие во всех регионах мира, где индоевропейские языки были в древности, от побережья Атлантики в Европе до Иранского нагорья, в том числе почти везде в Европе (за исключением Финляндии, Боснии и Герцеговины), Анатолии, Армении, Южной Сибири, во многих местах в Центральной Азии (в частности, СУАР, Туркменистан, Таджикистан и Афганистан), а также в Иране, Пакистане. В то время как R1b1 найдено в таком месте, как Левант и Камерун, R1b1b скорее всего возник в восточной Анатолии. |
Гаплогруппа – сестра основной русской гаплогруппы R1a. Родоначальник R1b родился около 15 000 лет назад в Центральной Азии. Примерно 10 000 лет назад род распался на три группы племен, которые начали расходиться в разных направлениях. Восточную ветвь рода ученые связывают с древними тохарами, от которых произошли современные уйгуры. Продвижение R1b на запад по-началу представляли одним потоком через Южный Урал и Русскую равнину в Западную Европу, потом начали появляться данные, не вписывающиеся в эту картину. Российский ученый Анатолий Клесов предложил гипотезу о том, что в районе Южного Урала отделилась группа племен, которая пошла на юг, преодолела Кавказ, территорию современной Турции, Египет, и по Североафриканскому побережью подошла к Гибралтарскому проливу, через который переправилась в Испанию. В районе современной франко-испанской границы эти два потока R1b встретились. Весной 2010 года эта гипотеза блестяще подтвердилась. Египетские ученые провели анализ мумии Тутанхамона, к их удивлению фараон оказался представителем рода R1b, но для российских ДНК-генеалогов это открытие было ожидаемым. Сейчас основная масса представителей рода проживает в Западной Европе, где R1b является основной гаплогруппой. В России только башкирский народ произошел от этой гаплогруппы. В русском народе R1b составляет 5%. В Петровскую и Екатерининскую эпоху проводилась государственная политика массового привлечения иностранных специалистов из Европы [в основном, немцев], современные русские R1b являются их потомками. |
Гаплогруппа R1b продвигалась из Южной Сибири, где образовалась 16 тысяч лет назад [и оттуда попала к тюркоязычным народам],
Путь в континентальную Европу с Пиренеев – это путь и времена культуры колоколовидных кубков, принадлежащей предкам пракельтов и праиталиков.
Я бы скорректировал первую часть этой версии. Гаплогруппа R1b сформировалась не в Южной Сибири, а на Южном Урале, где была одомашнена лошадь [что и стало причиной и возможностью такой масштабной миграции]. Именно оттуда коневоды - носители этой гаплогруппы - мигрировали 2 потоками на запад (в Причерноморье и на Северный Кавказ) и на восток (через Северный Казахстан в Южную Сибирь) [впрочем, могли и из Ю.Сибири через Ю.Урал, где познакомились с домашней лошадью]. Из Причерноморья тоже было 2 потока миграции - вверх по Дунаю и его притокам на север Западной Европы, и на Иберийский полуостров - возможно, морем (либо как описано - через Переднюю Азию и Северную Африку).
Это были строители мегалитов. А вот откуда они взяли эту традицию? Либо сами создали на Южном Урале или в Южной Сибири (Аркаим), либо переняли или создали на северо-запраде Кавказа (Адыгея). Оттуда эта традиция распространилась по побережью средиземного моря - вероятно, морским путем. И это были не языковые предки итало-кельтов, а биологические.
Миграция самих италиков и кельтов, как и распространение железа, произошло после Троянской войны (в начале I тысячелетия до н.э.), хотя, конечно, на этих территориях могли проживать их более древние родственные племена (как древние греки - осваивали Грецию несколькими волнами). Можно предположить, что народ колоколовидных кубков из Южной Сибири по языку был близок финно-угорским племенам, а, может быть, это были предки этрусков или даже древние енисейцы (или родственные им предки басков, жившие на Южном Урале, чей язык занимает как раз промежуточное положение между енисейскими и северо-кавказскими).
Хетты (2000-1200 до н.э.) были первыми индоевропейцами сделавшими вызов (и потерпевшими поражение) с могучей месопотамской и египетской империями. Доказано, что хеттский правящий класс был ответвлением от Майкопской культуры, который завоевал царство хаттов. Северная Анатолия, возможно, также была заселена индоевропейцами, которые позже основали Майкопскую культуру и распространили свой язык и культуру в Понтийско-Каспийской степи. Какой бы не был путь, северные анатолийцы бронзового века - это индоевропейские мигранты, и несомненно принадлежали по большей части к гаплогруппе R1b1a2. Хатты [?], возможно, были из некоторых старых ближневосточных R1b, смешавшихся с другими гаплогруппами, распространенными в Анатолии в настоящее время (G2a, J2a и E-M78).
Легендарная Троя вполне могла быть индоевропейской колонией для обеспечения торговых путей между Черным морем и Эгейским. Троянцы были лувийским перевалочным пунктом, связанным с хеттами (отсюда индоевропейские топонимы), с доказанными культурными связями с культурой Понтийско-Каспийской степи. Первые упоминания о Трое восходят к 3000 г. до н.э., в самом развитии Майкопской культуры, и во время производства первых кораблей. Учитывая раннее основание Трои, скорее всего, из двух индоевропейских гаплогрупп это были R1b1a2, а не R1a1a1.
Греческие R1b входят во многие разновидности: R1b1 с Ближнего Востока, R1b1b из Анатолии, и Европейские R1b1b2, в том числе прото-кельтские S116/P312 и Гальштат-кельтские S28/U152.
Наличие R1b1b2 в Греции может быть связано с вторжением Дориан, которое, как полагают, произошло в 12 веке до нашей эры. Дорийцы возможно были связаны с троянцами и хеттами, принадлежащими к древнейшим индоевропейским языковым группам, или прото-кельтами из Центральной Европы и долины Дуная. Среди их Y-ДНК линий были преимущественно R1b1b или R1b1b2. Дорийцы могут быть потомками первых (R1b) степных кочевников, которые обосновались в восточной части Балканского региона (культуры Чернавода и Эзеро) и не продолжили миграции вверх по Дунаю в Центральную и Западную Европу.
Греческие и анатолийские R1b-S28 линии могут быть связаны с кельтским вторжения в 3 веке до н.э., но более вероятно, с римской оккупацией. Старые клады из R1b, такие как R1b1 или R1b1a лишь в незначительном количестве и пришли с J2 и E1b с Ближнего Востока. Микенцы могли бы принести R1b1b2 в Грецию, но их происхождение можно проследить до культуры Сейсмино-Турбинского феномена северной лесостепи, которыми были в большей степени племена из N1c1 и R1a1.
Современные лингвисты поместили прото-индоевропейскую прародину в Понтийско-Каспийские степи - различные географические и археологические области от устья Дуная до Уральских гор и Северного Кавказа. Неолит, энеолит и ранний бронзовый века в культуре Понтийско-Каспийской степи были названы, согласно гипотезе Марии Гимбутас Курганной культурой (7000-2200 до н.э.), благодаря практике захоронения умерших под курганами между последовательностью культур в этом регионе ("курган I" - ...). Лошади были впервые одомашненные около 4000 гг. до н.э. (у других = 6000 лет до н.э. или даже раннее) в степи, возможно, где-то в районе Дона и Нижней Волги, и вскоре стали определяющим элементом степной культуры. Во времена бронзового века - периода, известного как горизонт Ямной Культуры (3300-2500 до н.э.), пастухи крупного рогатого скота и овец использовали повозки для транспортировки продовольствия и палаток, которые позволяли им двигаться дальше в степь, порождая новый мобильный образ жизни, что привело потом к большим индоевропейским миграциям.
Понтийско-Каспийские культуры степей можно разделить на западную часть, начиная от Дона до Днестра (а позже Дуная), и восточную - в Волго-Уральском регионе. Понтийские степи, вероятно, населенные мужчинами смешанных R1a и R1b линий, с более высокой плотностью R1b к северу от Кавказа, и R1a в северной степи и лесостепи. R1b почти наверняка перешли из Северной Анатолии в понтийско-Каспийской степи. Пока до конца не ясно, произошло это до, во время или после неолита. Регулярный поток R1b на Кавказе не может быть исключен. Генетическое разнообразие R1b больше всего в Закавказье, поэтому трудно отрицать, что R1b поселились и развивались там перед входом в степи мира. Означает ли это, что индоевропейские языки возникли в степи у популяций R1a и R1b, и что иммигранты слились в рассматривемой культуре? Или что прото-индо-европейский язык появляется в северной Анатолии или в Закавказье, а уже затем распространился в степи с R1b? Или же прото-индо-европейцы впервые появляются в степи, как гибридный язык Кавказских / анатолийских R1b и R1a? Этот вопрос не имеет очевидного ответа, но, основываясь на древности и архаичности анатолийских языков (хеттский, палайский, лувийский, лидийский, и так далее), гипотеза происхождения прото-индо-европейского языка в Анатолии заслуживает доверия. Кроме того, есть документальные подтверждения слов, заимствованных из кавказских языков в индо-европейских языках. Это намного более вероятно, произошло, если бы прото-индо-европейский развился около Кавказа, чем в далекой степи. За R1b, следовательно, было распространение прото-индоевропейских языков в степи, а оттуда в Европу, Центральную Азию и Юго-Восточной Азии.
Майкопская культура и гаплогруппа R1b ведут в степи? Майкопская культура (3700-2500 до н.э.) на Северном Кавказе, была культурным расширением Ямной культуры. Несмотря на это, как правило, она не считается частью понтийско-каспийской степной культуры благодаря своей географии, но на Северном Кавказе была тесно связана со степью, о чем свидетельствуют многочисленные керамика, золото, медь и бронза. Оружие и ювелирные изделия в современных культурах: Михайловка, Средний Стог Оба и Кеми. Связь между Северным Причерноморьем и Северным Кавказом старше Майкопского периода. Его предшественник, следующая культура (4400-3700 до н.э.), уже была в Суворово-Новоданиловке и в начале культуры Средний Стог, и даже старше аналогичной культуры Хвалынск на Волге. Это может быть период, когда R1b начали взаимодействие и смешение с R1a населением степей.
Люди Ямной и Майкопской культуры использовали курганные захоронения с их умершими в положении лежа с поднятыми коленями и ориентироваными по северо-восточной/юго-западной оси. Могилы были посыпаны красной охрой на полу а жертвенное животное хоронили рядом с людьми. Они также знали верховую езду, телеги, крупный рогатый скот и овец, использовали в их экономике, основанной на использовании медного / бронзового боевых топоров (в качестве молотов) и кинжалов. В самом деле, старые телеги и бронзовые артефакты находятся на Северном Кавказе, а уже оттуда распространялись по степи. Майкоп был передовой культурой эпохи бронзы, на самом деле это одна из первых археологических культур, принявших участие в развитии металлообработки, и, следовательно, оружия из металла. Стариный меч в мире был найден в конце Майкопской культуры в залежах кургана 31. Его стиль напоминает длинный кельтский меч. Кости лошадей и изображения лошадей появляются уже в начале Майкопских могильников, предполагая, что в культуре Майкопа, возможно, сформировался степной тип людей, которые имели тесные связи с ними. Тем не менее, присутствие элементов культуры коренным образом отличается от степной культуры и в некоторых местах может означать, что Майкоп был гибридным по населению. Без анализа ДНК нельзя говорить, были ли эти две группы населения анатолийской R1b и кавказской G2a, или носители гаплогруппы R1a, которые поселились здесь позже. Две или три народности, возможно, даже сожительствовали бок о бок в различных населенных пунктах. Типичная кавказская Y-ДНК линия (например, G2a) не последовала примеру индоевропейских миграций, поэтому смешанные браки должны были быть ограничены, по крайней мере, ограниченны для индо-европейских мужчин с кавказскими женщинами, а не наоборот.
Майкопские люди являются теми, кто ввел внедрение примитивных колесных транспортных средств (телег) от Месопотамии до степей. Это был революционный способ жизни в степи, а в дальнейшем привел к развитию (конных) боевых колесниц около 2000 года до нашей эры. Кавалерия и колесницы играют важную роль в последующих индоевропейских миграциях, что позволяет им быстро и легко победить любого с кем они столкнулись. В сочетании с современным оружием и бронзовыми технологиями, западная ветвь (R1b) индо-европейцев от берегов Черного моря являются превосходными кандидатами на то, что это таинственный народ моря, который ворвался к восточному берегу Средиземного моря в период второго тысячелетия до нашей эры. Распространение индо-европейского языка хеттов в центральной Анатолии произошло спустя несколько веков после исчезновения Майкопской культуры. Обратный переход от Северного Кавказа до северной Анатолии, очень вероятно, произошел в этот периоде экспансии. Не вызывает сомнений то, что хетты использовали колесницы, изобретенные в Волго-Уральской степи. R1a были обнаружены с низкой частотой в Армении и Северной Анатолии, что дает основания предположить, что гибридные группы R1a, R1b из Волго-Уральского региона мигрировали в этот регион в промежутке между 2000 г до н.э. и 1650 до н.э.
Майкопскую и Ямную культуры сменила Срубная культура (1600-1200 до н.э.), возможно, представленная из носителей гаплогруппы R1a1 северных и восточных степей, распространившихся к берегам Черного моря.
На заре цивилизаций группы R1b людей, как полагают, мигрировали из региона Каспийского моря в Центральную Азию, где она превратилась в R1b1b1 (M73) субклад. Эта разновидность R1b встречается почти исключительно в очень специфических местах у населения Центральной Азии. Самый высокий процент обнаружен среди уйгуров Синьцзяна (до 20%) на северо-западе Китая, у хазарейцев народа Афганистана (до 30%) и у башкиров Абзелиловского района (до 43%) в России (граница с Казахстаном).
В Центральной Азии R1b1b1 может соответствовать тохарской ветви индоевропейцев. Вполне возможно, что тохары отделились от основной линии R1b в начале 7000 г. до н.э. На протяжении столетий некоторые группы этих кочевых племен мигрировали около Южного Урала, другие возле бассейна реки Тарим (Синьцзян) и в южной части Центральной Азии. Другая теория состоит в том, что группа ранних наездников [тохары?] из культуры Репина (3700-3300 до н.э.) мигрировали из Дон-Поволжья в горы Алтая, основав культуру Афанасьево (ок. 3600-2400 до н.э.), а затем переместились на юг в Тарим.
Мумии светловолосых людей европеоидной расы были найдены в бассейне Тарим, самые старые из которых датируются 1800 г. до нашей эры. Современные жители бассейна реки Тарим, уйгуры, относятся к тому же R1b-M73 субкладу (до 20%) и R1a1 (около 30%). Это может означать, что они сформировались из гибридных R1b и R1a обществ к тому времени и достигли бассейна реки Тарим. Но R1a1 могли бы также прибыть самостоятельно в течение более поздних миграций культур Андроновского круга (ок. 2000 до н.э.), или гораздо позже, через несколько кочевых скифо-иранских или тюркских племен (после 700 г. до н.э.)
Наличие у Иидоевропейцев бронзового оружия и лошадей дали им огромное преимущество перед коренными обитателями Европы, а именно перед гаплогруппой I (потомки первых европейских кроманьонцев), и первых неолитических культур пастухов и фермеров (G2a, J2, E-V13 и T). Это позволило гаплогруппам R1a и R1b заменить большинство старых мужских линий в Западной Европе, хотя женские линии, кажется, были менее затронуты. По сравнению с индо-иранским вторжением в Южную Азию показательно, что около 40% мужских линий северной Индии принадлежат гаплогруппе R1a, но менее 10% женских линий может быть названо индо-европейскими по происхождению. Влияние индоевропейцев было более тяжелым в Европе, потому что европейское общество 4000 лет назад было менее развито с точки зрения сельского хозяйства, технологии (без бронзового оружия) и плотность населения ниже, чем в цивилизациях долины Инда. Это особенно справедливо в западноевропейских странах, где сельское хозяйство пришло намного позже, чем на Балканы и в Центральную Европу. Греция, в целом Балканы и Карпаты - были самые передовые европейские страны в то время и меньше всего пострадали в плане замены гаплогрупп. Местные европейские Y-ДНК гаплогруппы (I1, I2a, I2b) также сохранились лучше в тех регионах, которые были более труднодоступны и менее гостеприимны - как в Скандинавии, Бретании, Сардинии или в Динарских Альпах. Первые нападения степных людей на Балканы произошли между 4200 до н.э. и 3900 до н.э., когда наездники перешли Днестр и Дунай и, видимо, разрушили города Варну и Караново культуры VI в Восточной Румынии и Болгарии. Изменение климата в результате холодных зим в течение этого периода, вероятно, подтолкнуло степных пастухов искать мягкие пастбища для своего скота, что привело к голоду и внутренним беспорядкам в дунайской и балканской общинах. Последовавшая за этим Чернаводова культура (4000-3200 до н.э.) и Эзеро культура (3300-2700 до н.э.), кажется, были из смешанного населения степных иммигрантов и людей из старых поселений. Эти степные иммигранты, вероятно имели смесь обоих гаплогрупп R1a1 и R1b. Многие дунайские фермеры также мигрировали в Кукутень-Триполье в Восточных Карпатах, в результате чего население направилось на северо-восток расширяясь до долины Днепра, в результате чего Y-гаплогруппы E-V13, J2b и T есть в настоящее время в центральной Украине. Это преждевременное движение индоевропейских народов на запад было довольно ограниченным, в связи с отсутствием бронзового оружия и организованной армии в то время, и было действительно возможно только благодаря климатическим катастрофам. Карпатская, дунайская и балканская культуры были слишком густонаселенными и технологически продвинутыми, чтобы обеспечить массовую миграцию. Бронзовый век анонсирует новое развитие. Носители гаплогруппы R1a1, вероятно, были первыми, кто успешно проникнул в самое сердце Европы, в составе культуры шнуровой керамики (3200-1800 до н.э.) как естественная экспансия на запад Ямной культуры. Они пошли на запад, в Германию и Скандинавию. Анализ ДНК из продовольственной культуры Иелау подтверждает наличие гаплогруппы R1a1 в центральной Германии около 2600 до нашей эры. Эта культура вполне могла бы расширится в лесостепи, или на северной окраине Ямной культуры, где были распространены R1a линии. Гаплогруппа R1b1a2, как полагают, прибыли в Центральную и Западную Европу около 3000 г. до н.э., идя вверх по Дунаю от побережья Черного моря, другой поток через Анатолию и Кавказ и третий, возможно, через северную Африку на Пиренеи. Археологические и генетические данные (распределение субклада R1b), указывают на несколько последовательных волн в направлении Дуная между 2800 до н.э. и 2300 до н.э. (начало Унитецкой культуры). Интересно отметить, что это также соответствует концу культуры Майкопа (2500 до н.э.) и Обской культуры (2200 г. до н.э.) на северном побережье Черного моря, где замена культур происходит из миграций со стороны северных степей. Поэтому может быть предположено, что (в основном) R1b население из северной части Черного моря мигрировали на запад из-за давления со стороны других индоевропейских народов (R1a) с севера, в виде растущей прото-индо-иранской ветви, связанной с современными Абашевской и Полтавской культурами. Возможно, что культура Колоколовидных Кубков (2800-1900 до н.э.) была уже индоевропейской (хотя они были под влиянием культуры шнуровой керамики), потому что они были в непосредственной связи с родной мегалитической культурой. Есть вероятность, что кубки и лошади обнаруженные в Западной Европе в этот период были результатом торговли с соседними индоевропейскими культурами, в том числе первой волны R1b в Центральной Европе. Тем не менее, нельзя отрицать, что следующие Унитецкая культура (2300-1600 до н.э.), культура Курганов (1600-1200 до н.э.), Культура полей погребальных урн (1300-1200 до н.э.) и Гальштатская (1200-750) культуры были связаны с распространением R1b в Европе, - они резко внедряют новые технологии и внедряют совершенно иной образ жизни. Действительно ли индо-европейцы вторглись в Западную Европу? Сторонники модели преемственности палеолита или неолита утверждают, что бронзовые технологии и лошадей можно было бы импортировать у западных европейцев от их восточноевропейских соседей, и что фактически индоевропейского вторжения могло не быть. Труднее понять, как Италийские, кельтские и германские языки были приняты Западной и Северной Европой без хотя бы небольшого вторжения. Было высказано предположение, что индоевропейская ветвь языков просто распространяется через контакт, как и технологии, или потому, что он был языком небольшой элиты и, следовательно, его принятие воспринимается как престижное. Однако людям не просто изменить язык, потому что это звучит лучше, или более престижно. Языковой разрыв между пред-индо-европейскими наречиями был примерно таким же большим, как между современным английским и китайским языками. С лингвистической точки зрения, только широкомасштабной миграцией носителей языка можно объяснить массовое принятие индо-европейских языков в Западной Европе - оставляя только басков как пережиток языков неолита. Одним из важных археологических аргументов в пользу замены неолитических культур индо-европейской культурой в эпоху бронзового века исходит из керамических стилей. Внезапное появление бронзовой технологии в Западной Европе совпадает с керамикой и неожиданно становится все более простым и менее украшенным, как и в понтийских степях. До тех пор гончарные изделия были постоянно в эволюционировании в сторону большей сложности и деталей более чем 3000 лет. Люди не просто решили вернуться к более примитивным стилям. Возможно, одно изолированное племя могло экспериментировать с чем-нибудь попроще, но каковы шансы, что далекие культуры Иберии, Галлии, Италии и Великобритании все решили провести такой невероятный сдвиг примерно в то же время? Лучшее объяснение, что этот новый стиль был введен иностранными захватчиками. В данном случае это не просто спекуляции, есть достаточно доказательств, что это простая керамика характерно связана с появлением прото-индо-европейского языка. Кроме керамики, археология дает достаточно доказательств, что ранний бронзовый век в Центральной и Западной Европе совпадает с радикальным сдвигом в производстве продуктов питания. Сельское хозяйство испытывает резкое сокращение в обмен на повышенное внимание в одомашнивании скота. Это также период, когда лошади все чаще используются и коровье молоко потребляется регулярно. Даже после введения сельского хозяйства около 5200 до н.э., Буго-днестровская культура и поздние культуры степей характеризуются в экономике доминированием животноводства, только с ограниченным сельским хозяйством. Эта модель расширяет свою деятельность в Европе, именно в то же время, как появляется бронза. Религиозные верования и искусство прошли полный переворот в бронзовом веке в Европе. Неолитические общества на Ближнем Востоке и в Европе всегда поклонялись женским фигуркам в форме культа плодородия. Степные культуры, наоборот, не производили женских фигурок. С распространением бронзовых технологий в долине Дуная в Западной Европе, символы плодородия и плодовитости постепенно исчезают и заменяются скульптурами домашних животных. Другим признаком того, что индо-европейские народы пришли в большом количестве в регион Центральной и Западной Европы можно найти в практике захоронений. Европейцы неолита или кремировали своих умерших (например, Кукутень-трипольская культура) или хоронили их в коллективных могилах (это было в случае мегалитических культур). В степи каждый человек был похоронен отдельно, и высокопоставленные могилы помещали в погребальную камеру, увенчанную круговой насыпью. Обычно клали оружие (булавы, топоры, кинжалы), лошади костей, и демонтированные телеги (или более поздние колесницы). Эти характерные курганы известны как курганы в понтийских степях. Мужчины получили больше роскошных гробниц, чем женщины, даже среди детей, и различия в иерархии очевидны между захоронениями. Индоевропейцы были сильно иерархическим мужским обществом, в отличие от более равноправного и матрилинейного общества культур Старой Европы. Распространение статуса доминирующих мужчин в Центральной Европе в эпоху бронзы является явным признаком, что правящая элита теперь стала индоевропейской. Эта практика также распространяться в Центральной Азии и Южной Сибири - двух регионах, где R1a и R1b линии находятся в настоящее время, как в Центральной Европе. Церемония захоронения - один из самых эмоциональных и личных аспектов культуры. Весьма сомнительно, что люди будут менять своих предков практикой "просто сделать, как у соседей". На самом деле, различные погребальные методы сосуществовали бок о бок во время европейского неолита и энеолита. Принятие некоторых элементов иностранной культуры как правило, происходит, если одна цивилизация показывает для соседних культур свое превосходство. Этот процесс называется "аккультураций". Однако ничто не указывает, что степная культура была настолько культурно превосходной, чтобы мотивировать весь континент, даже Атлантические культуры на протяжении 2000 км от понтийских степей, к оставлению многих таких фундаментальных символов собственной древней культуры, и даже свой собственный язык. На самом деле, старая Европа была гораздо более изысканной в своей керамике и ювелирных изделиях, чем "грубые" люди степи. У Индо-европейского первенства были культурные преимущества, а также военные, благодаря лошадям, бронзовому оружию и нормы профессиональной оценки отдельных героических подвигов во время войны (эти этические ценности, как известно из старых текстов, таких как Ригведа, Авеста, или микенской и Хеттской литературы). После лингвистики и археологии, третья категория доказательств исходит от генетики. Впервые было предположено, что R1b - родом из Западной Европы, потому что это место, где она была наиболее распространенной. С тех пор было доказано, что гаплотипы гаплогруппы R1b отображаются выше микросотелитных разнообразий в Анатолии и на Ближнем Востоке, чем в Европе. Европейские субклады также более поздние, чем на Ближнем Востоке и в Центральной Азии. Основные европейские субклады, R-P312/S116 только восходят от 3500 до 3000 до нашей эры. Это не значит, что самые старые общие предки этой линии прибыли в Западную Европу в этот период, но это первый человек, который нес мутацию R-P312/S116 жил по крайней мере 5000 лет назад, предположительно где-то около Черного моря. В любом случае этот срок слишком поздний для происхождения из палеолита, мезолита или даже неолита для R1b в Европе. Открытие того, что считалось «Европейской линией» в Центральной Азии, Пакистане и Иране вбило "последний гвоздь в гроб" происхождения палеолитических R1b в Западной Европе, и подтвердило индоевропейские гипотезы. Все элементы сходятся в пользу крупномасштабной миграции на основе верховой езды индоевропейских народов в Западной Европе [или в в Западную Европу?] между 2500 до 2100 до н.э., что способствует смене образа жизни неолита и энеолита - по сути новая культура бронзового века, с простой глиняной посудой, земледелием, скотоводством приобретает новые ритуалы и новые значения (для мужской линии общества, воинов и героев), которые не развиваются из местных предшественников. Эти прото-итало-кельто-германские R1b люди поселились вокруг Альп 2300 году до н.э., и, судя по распространению бронзовых работ, достигли Иберии ппримерно в 2250 до н.э., Великобритании к 2100 году до нашей эры и Ирландии в 2000 году до нашей эры. Это первая волна R1b предположительно осуществлялась R1b-L21 линией в большем количестве, так как они встречаются повсеместно в западной, северной и центральной Европе. Второе расширение R1b состоялось с культурой полей погребальных урн / Гальштатской культуры около 1200 г. до н.э., направляясь на запад до Атлантического океана, к северу до Скандинавии, и на восток до Греции и Анатолии. Новая культура бронзового века процветала вокруг Альп (от Унитецкой до начала Гальштатской) благодаря обилию металла в регионе, а также заложены основы для классической кельтской культуры. Альпийские кельты Гальштатской культуры, связанные с S28 (ака U152) мутацией, отделились и (в том числе S28/U152) вступили в Италию в 1200 г. до н.э., но было, конечно, несколько последовательных волн, о чем свидетельствуют более поздние прибытия Цизальпинских кельтов. Белги были также S28/U152, расширение культуры - Латенская на север, дальше за Рейн, Мозель, Мез и впадающие реки. Одна из распространенных языковых черт в галльских / бриттских и кельтских языках, связанных с расширением Гальштатской культуры указывает, что они поменяли отдельные звуки. Они, как известно лингвистам, считаются P-кельтской ветвью. Считается, что это изменение произошло в связи с неспособностью произносить звук, естественный для индоевропейского населения Центральной Европы, Галлии и Италии, которые были носителями афро-азиатских диалектов, которые произошли от ближневосточных языков. Этруски, хоть и более поздние пришельцы из Леванта, также вписываются в эту категорию. Недавно была высказано гипотеза, что кельтские языки заимствовали часть своей грамматики из афро-азиатских языков [препозиционное сказуемое]. Это изменение могло произойти, когда прото-итало-кельты перешли из степи в бассейне Дуная и смешались с населением ближневосточных фермеров принадлежащих гаплогруппам E-V13, T, G2a и J2b. Однако такой ранний переход не может объяснить, почему слова кельтского языка развились в Ирландии и Иберии. Более вероятно, что смешение произошло после того, как итало-кельты впервые распространились во всех странах Западной Европы. R1b-S28/U152 в связи с P-кельтским означает, что произошел переход вокруг Альп и Италии после 1200 г. до нашей эры [после Троянской войны]. R1b-S21 (ака U106) найден в высоких концентрациях в Нидерландах и Северной Германии. Его присутствие в других странах Европы могут быть отнесены к 5-й и 6-м векам германских миграций. Фризы и саксы распространили эту гаплогруппу на Британских островах, франки в Бельгии и Франции, а лангобарды в Австрии и Северной Италии. Высокая концентрация S21/U106 вокруг Австрии могла возникнуть там, в Гальштатский период, или возникла вокруг Черного моря и перешла туда в Гальштатский период. На самом деле, на юге Германии и Австрии, вместе взятые имеют самое высокое разнообразие R1b в Европе. Кроме S21, три важнейших первых субклада уровня R1b1b2a1b (L21, S28, M167) находятся в этом районе в разумных частотах, чтобы усмотреть распространение из Унитецкой в Гальштатскую культуру с остальной Западной Европой. Как R1b пришли на смену большинству старых линий в Западной Европе? До недавнего времени считалось, что R1b зародилась в Западной Европе из-за своего максимального присутствия в регионе и по сей день. Считалось, что R1b - представители палеолита Европы (европейские кроманьонцы), которые нашли убежище в франко-Кантабрийском регионе на пике последнего ледникового периода, то реколонизация Центральной и Северной Европы после отступления ледяного покрова. Филогенией R1b доказано, что этот сценарий не существовал, потому что старшие клады R1b последовательны в Центральной Азии и на Ближнем Востоке, а более молодые в Западной и Северной Европе. Существовал четкий градиент с востока на запад отслеживания миграции R1b. В этом время основной переход от берегов Черного моря до Центральной Европы также произошел в соответствии со сроками индоевропейского вторжения в Европу, которые совпадают с появлением культур бронзового века в Западной Европе, и распространением итало-кельтских и германских языков. Историки и археологи долгое время утверждали была ли индоевропейская миграция массированным вторжением, или, скорее, культурная диффузия языка и технологии распространялись только на небольшое количество пришельцев. Ответ на этот вопрос вполне может быть "нет". Сторонники теории диффузии хотели бы заставить нас думать, что R1b является родным субкладом для Западной Европы, а только R1a представлять индоевропейцев. Проблема в том, что гаплогруппа R возникает в Центральной Азии или Южной Азии, и R2 по-прежнему ограничено Центральной и Южной Азии, в то время как обнаруженные древние линии R1a в Европе, а старшие субклады из R1b - в Азии. Возраст субклада R1b в Европе совпадает с бронзовым веком. R1b, следовательно, должна заменить большинство местных Y-ДНК линий в Европе с бронзового века и позже. Тем не менее, массовая миграция и почти полная аннигиляция населения палеолита вряд ли может быть предусмотрена. Западные европейцы есть в Ирландии, Голландии, Португалии или Аквитании, несмотря на все регионы, где R1b является доминирующим. Аутосомные исследования ДНК подтвердили, что западноевропейское население далеко не однородно. Много материнских линий (мтДНК) также, вероятно, происхождением из палеолита (например, H1, H3, U5 и V), основанные на древних ДНК. То, что многие люди забывают о том, что есть и не нуждается в крупномасштабных исходах для мужской линии заменить достаточно быстро. И вот почему. Многоженство. В отличие от женщин, мужчины не ограничены в количестве детей, они могут производить потомство. Мужчины как правило, имеют больше детей. Это было справедливо в примитивных обществах, где полигамия часто была нормой для вождей и королей. Статус и питание. Оснащенные бронзовым оружием и лошадьми, индоевропейцы легко покорили неолитических фермеров и потом с еще большей легкостью охотников-собирателей Европы. Если они не истребляли коренных европейских мужчин, новички стали бы новым правящим классом, с Множеством местных царей, вождей и знати (кельты и германцы жили в маленьких сельских общинах с вождем, отдельная часть небольшого племени во главе с королем) с более высокими репродуктивными возможностями, чем в среднем. Вторжение армии как правило, имеют гораздо больше мужчин, чем женщин. Люди должны поэтому найти женщин в покоренном населении. Войны ведутся людьми, и проигравшие имеют более тяжелые потери, в результате чего все большее число женщин попадает в распоряжение победителей. Индоевропейцы были воинственным народом с сильной героической смелостью, подчеркивая честь и воинские доблести. Их превосходная технология (металлическое оружие, транспортные средства и герои) и отношение к жизни позволило бы им победить любую группу населения, которая не имеет организованной армии с металлическим оружием (то есть никому, кроме ближневосточных цивилизаций). Есть ли генетическая предрасположенность к зачатию мальчиков у R1b? Основная роль Y-хромосомы в теле человека состоит в запуске развития организма по мужскому типу. Гаплогруппы определяются на основе дифференциации мутации Y-хромосомы в основном в негенных областях. Каждая мутация теоретически способна отрицательно повлиять на производство спермы и подвижность сперматозоидов, если она выпала на участок с геном, например SRY. Предварительные исследования уже установили связь между определенной мутацией в Y-гаплогруппе и увеличением или уменьшением подвижностью сперматозоидов, чаще всего эти мутации патологические и приводят мужскому бесплодию. Чем выше подвижность, тем выше шансы на зачатие мальчика. Абсолютно исключено, чтобы R1b или любая другая гаплогруппа древнее 5 000 лет, имела преимущество в сторону большего мужского потомства. Увеличение или уменьшение подвижности сперматозоидов вследствие мутаций скорее могут быть характерны для отдельных молодых групп в разных гаплогруппах и субкладах. В любом случае, если на начальном этапе в этносе по какой-либо причине будет рост носителей отдельной гаплогруппы, то скорее всего в последующем, в этой популяции сохраниться такая же пропорция - это называется эффектом основателя. Смещения существуют только по отношению к другим гаплогруппам и обнаружены в одной и той же популяции. Вполне возможно, что сравнительно недавно R1b субклад Западной Европы имели значительное преимущество по сравнению со старыми гаплогруппами в этом регионе, в частности гаплогруппами I2 и G2a. Замена мужской линии по этой модели быстро становится экспоненциальной. Представьте себе 100 индо-европейских мужчин завоевывает 1000 коренных европейцев (соотношение 1:10). Военные потери привели к более высокой доли женщин в покоренном населении. Давайте предположим, что уцелевшее население состоит из 700 женщин и 300 мужчин. Давайте предположим, что победоносные индо-европейские мужчины в конечном итоге, имеют в два раза больше детей, достигнув зрелого возраста, чем мужчины побежденного племени. Существует ряд причин для этого. Победители займут больше жен, наложниц или даже изнасилуют женщин побежденного племени. Их высокий статус будет гарантировать их больше богатства и, следовательно, более качественное питание для своего потомства, увеличивая шансы на достижение совершеннолетия и продолжения своего рода. Потомства в соотношении 2 к 1 для мужчин на самом деле является консервативной оценкой, так как это полностью возможно, что в бронзовом веке чувства привели бы к уничтожению большинства мужчин на проигравшей стороне (как свидетельствует Ветхий Завет). Тем не менее, это займет всего несколько поколений на победу Y-ДНК линий, чтобы стать большинством. Например, если в первом поколении индоевропейцев было два оставшихся в живых сына на человека, в отношении только одного человека на коренных жителей, число индоевропейских отцовских линий будет проходить в 200 человек во втором поколении, 400 в третьем, 800 в четвертом и 1600 в пятом, и так далее. За это время коренным линиям грозит только стагнация на 300 человек для каждого поколения. На основе такого сценария, R1b линии быстро подавили бы местные линии. Даже если у индоевропейских завоевателей было лишь немного больше детей, чем у местных мужчин, R1b линия станет доминирующей в течение нескольких веков. Кельтская культура продолжалась более 1000 лет в континентальной Европе до римского завоевания, чтобы положить конец привилегии от вождей и знати. Это более чем достаточно по времени для R1b линий достичь от 50 до 80% населения. Современные частоты R1b формируют градиент по краю Атлантической Европы (высокий процент) в Центральной и Восточной Европе (самый низкий), вновь поднимается на анатолийской родине. Это почти наверняка связано с сельским хозяйством, которое лучше развито в Восточной и Центральной Европе, а затем, с более высокой плотностью населения, в результате чего R1b вторженцев здесь меньше, чем на Западе. Кроме того, другие индо-европейские культуры шнуровой керамики (R1a) уже прошли путь от современных России и Украины на запад, в Германию и Скандинавию. Было бы трудно для R1b людей, чтобы конкурировать со своими двоюродными братьями R1a, кто разделяет аналогичную технологию и культуру. Предварительно кельто-германские R1b поэтому вынуждены были уходить дальше на запад, прежде всего в Альпы, то есть обгоняя, те малонаселенные части Западной Европы. |
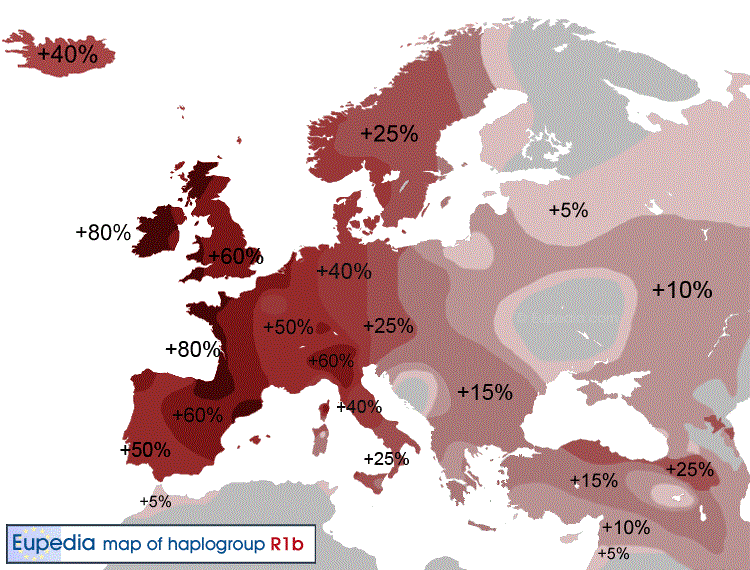
Современная концентрация (но не разнообразие) гаплогруппы R1b максимальна на территориях, связанных с кельтами и германцами: в южной Англии около 70 %, в северной и западной Англии, Испании, Франции, Уэльсе, Шотландии, Ирландии — до 90 % и более. Во всех этих странах распространен субклад R1b1a2 Также, например, у басков — 88,1 %, испанцев — 70 %, бельгийцев — 63 %, итальянцев — 40 %, немцев — 39 %, норвежцев — 25,9 % и других. У народов Восточной Европы встречается значительно реже. У осетин Алагира - 43 %, чехов и словаков — 35,6 %, поляков — 11,6 %-16,4 %, латышей — 15 %, венгров — 13,3 %, эстонцев — 9 %, литовцев — 5 %, беларусов — 4,2 %, русских — от 2,8 % до 21,3 %, украинцев — от 2 % до 18,9 %. На Балканах — у греков — от 13,5 % до 22,8 %, словенцев — 21 %, албанцев — 17,6 %, болгар — 17 %, хорватов — 15,7 %, румын — 13 %, сербов — 10,6 %, герцеговинцев — 3,6 %, боснийцев — 1,4 %. |
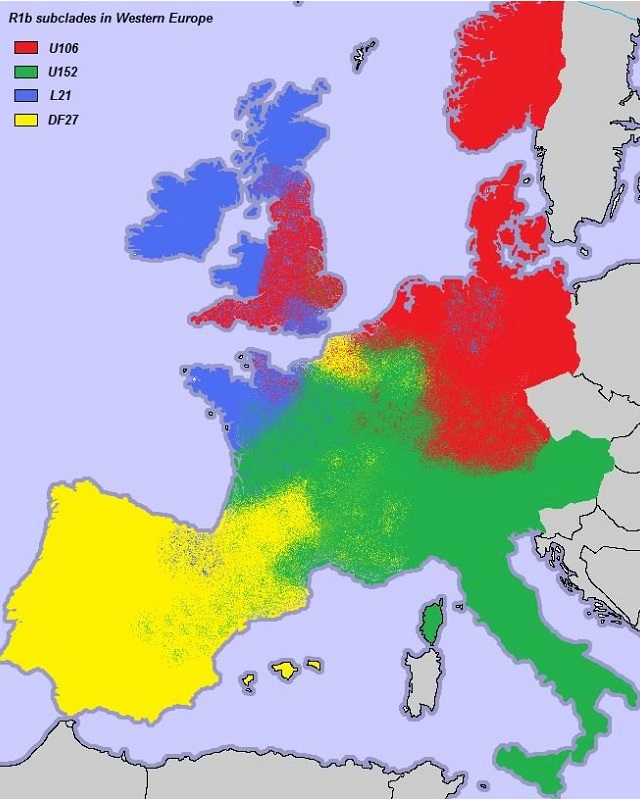
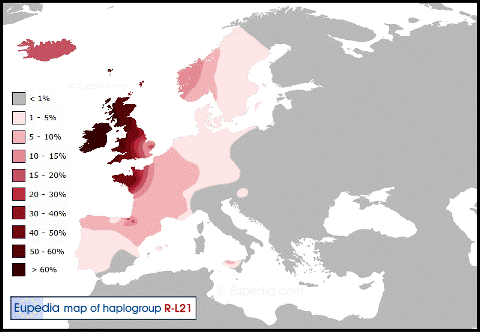
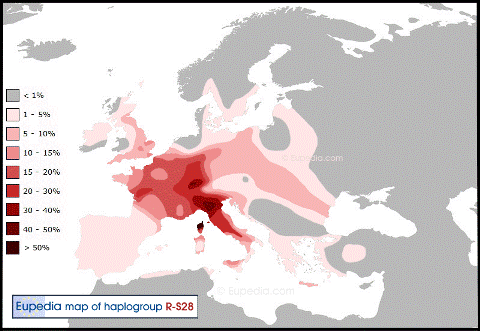
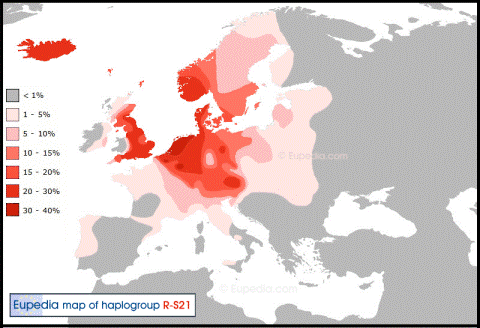
В западной Европе имеют распространение по большей части 3 основных субклада, миграции которых уже в Европе проходили, по большей части, независимо друг от друга. Это субклады L21, S28/U152 и S21/U106.
|
|
|
|
Субклад L21 можно условно назвать северо-западным или кельтским.
Его основной ареал приходится на Ирландию, Уэльс, Шотландию и французскую Бретань.
|
Субклад S28 можно условно назвать альпийским или северо-итальянским.
Его основной ареал приходиться на Альпы, север Италии и французский остров Корсика.
|
Субклад S21 можно условно назвать германским. Его частота резко повышается
в тех странах, по территории которых в древности пролегали пути миграций германских племен.
Основной ареал приходиться на Норвегию, Исландию, Англию, Швецию, Германию и Нидерланды.
|
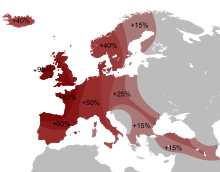
Современная концентрация R1b максимальна на территориях, связанных с кельтами: в южной Англии около 70 %, в северной и западной Англии, Испании, Франции, Уэльсе, Шотландии, Ирландии — до 90 % и более, в Испании — 70 %, в Бельгии - 63 %, во Франции — 60 %. По-видимому она связана с докельтским субстратом, поскольку высока её концентрация и у не-кельтов басков — 88,1 % и испанцев — 70 %. Кроме того, известно, что, например, строителями Стоунхенджа в Англии было население, обитавшее на острове до прихода кельтов. У соседних народов концентрация данной гаплогруппы падает: у итальянцев — 40 %[13], немцев — 39 %[14], норвежцев — 25,9 %[15] и других. У народов Восточной Европы встречается еще реже: У осетин Алагира - 43 %, чехов и словаков — 35,6 %, поляков — 11,6 %-16,4 %, латышей — 15 %, венгров — 13,3 %, эстонцев — 9 %, литовцев — 5 %, беларусов — 4,2 %, русских — от 2,8 % до 21,3 %, украинцев — от 2 % до 18,9 %. На Балканах: у греков — от 13,5 % до 22,8 %, словенцев — 21 %, албанцев — 17,6 %, болгар — 17 %, хорватов — 15,7 %, румын — 13 %, сербов — 10,6 %, герцеговинцев — 3,6 %, боснийцев — 1,4 %. Плотность распространения гаплогруппы R1b1a2 примерно совпадает с районами строительства мегалитов в Западной Европе. Это совпадение служит одним из оснований гипотезы Сайкса и Оппенгеймера об автохтонном палеоевропейском происхождении данной гаплогруппы. Согласно данной гипотезе, носители гаплогруппы R1b являются потомками Солютрейской культуры, пережившими последний ледниковый максимум в Западной Европе в изоляции от других народов, однако носителей предковой гаплогруппы R не могло быть среди людей, впервые заселивших Европу 40-30 тысячелетий назад, поскольку её древность оценивается всего в 30-25 тысячелетий и другие ветви группы R распространены в Восточной Европе и Центральной Азии, а потомки P и K распространены в максимальном разнообразии в Восточной Азии. Таким образом получается, что сразу после появления и распространения группы R часть её очень быстро переселилась на запад в Европу прямо посреди ледникового максимума, что выглядит невероятным. Ряд более новых исследований также подвергают критике гипотезу Сайкса. В частности, предполагается, что R1b следует ассоциировать с индоевропейцами (в частности в более позднее время в Западной Европе — особенно с кельтами), пришедшими из Причероморья примерно 5000 лет назад (предполагается, что изначально также носители праиндоевропейской культуры с гаплогруппой R1a занимали преимущественно северную часть ареала, тогда как R1b — более южную, возможно, включающую Кавказ и Анатолию). Культура Колоколовидных Кубков - предположительно является археологической культурой в составе которой гаплогруппа R1b1a2 заселяла Европу с берегов запада Северной Африки, но у современных народов населяющих этот регион гаплогруппа R1b1a2 составляет не более 5%. При этом возникает некоторая сложность объяснения того, почему большинство современных мужчин Западной Европы оказываются потомками выходцев из Понтийского региона. Однако приводятся демографические выкладки, показывающие, что в случае установления гегемонии со стороны пришельцев, технологически превосходивших автохтонное население, мужские линии победителей могут полностью вытеснить мужскую линию наследственности местного населения за несколько веков. Значительная часть мужчин могла быть убита сразу в военных столкновениях, в дальнейшем же семьи местных мужчин получали более низкий статус и могли воспроизводить меньше детей. Со времени установления в Западной Европе господства кельтов до более лояльных времён Римской империи прошло примерно восемь веков, за которые автохтонные гаплогруппы вполне могли полностью исчезнуть. В Центральной и Восточной Европе ко времени вторжения индоевропейцев существовала более развитая земледельческая культура, поэтому в этих районах вытеснение старых гаплогрупп не было таким полным. |
За пределами Западной Европы высокая концентрация данной гаплогруппы встречается лишь у некоторых популяций башкир (бурзян, тамьян, гайна и др.) — до 87%, что, возможно, связано с присутствием кроманьонского дотюркского субстрата. На Кавказе найдена у осетин-дигорцев — до 43 % и армян — 32,4 %. В Турции достигает (в среднем) 16,3 % (а у самих турок - 31%), Ираке — 11,3 %, также найдена и в других странах Западной Азии. В Центральной Азии обнаружена, в частности, у туркменов — 36,7 %, узбеков — 9,8 %, татар — 8,7 %, казахов — 5,6 %, уйгуров — от 8,2 % до 19,4 %. В Пакистане — 6,8 %. В Индии незначительна — 0,55 %.
Что интересно, R1b является доминирующей гаплогруппой у армян, и в районе Арцаха (Карабаха) она доходит до 40-50%, при этом отсутствуя (или присутствуя в малом количестве) у многих соседних народов. На основе этого появилась версия, что одним из очагов распространения этой гаплогруппы было Армянском Нагорье. Возможно, в Западную Европу носители этой гаплогруппы переселялись через Анатолию.
У бурушо (хунза), носителей языка бурушаски (и его вершикского диалекта), народа обитающего Пакистане и относящегося к дене-кавказской языковой общности, различные субклады R не просто заметно представлены, но преобладают, и резко преобладают. В том числе: R1a1—25 %, R2—14 %, R-М207—10 %, R1-М173—1 %, R1а1d-PK5—2 % (Firasat et al. 2007: 123). Как видим, совокупно гаплогруппа R у бурушо достигает 52%.
Особо обратим внимание здесь и на высокую частоту наиболее архаичной R-М207 — т. е., парагруппы R*, а также встреченную исключительно у бурушо R1а1d-PK5.
Но действительно адекватное представление о ситуации дает сопоставление частот субклад R у бурушо со средними частотами по Пакистану: для М207 — 0,6%, R2 — 8%, R1-М173 — 5%, и R1a1 — 37%.
В Северной Африке: у алжирских арабов из Орана — 10,8 %, тунисских арабов — 7 %, алжирских берберов — 5,8 %, в Марокко — около 2,5 %, у арабов Египта — менее 1 %.
В субсахарской Африке: в Камеруне - 95,1%.
Субклад R1 развился в R1b*, потом в R1b1* в северной части Ближнего Востока во время ледникового периода. Это произошло предположительно при переходе в северную Анатолию и на Кавказ в начале неолита, где появился субклад R1b1a. Ближневосточные остатки превратились в R1b1a (M18), который теперь встречается в низких частотах у ливанцев и у друзов. Финикийцы (которые пришли из современного Ливана) распространили эту гаплогруппу R1b1a и R1b1* в своих колониях, в частности, на Сардинии и в Магрибе.
Субклады R1b1a1 и R1b1a2 (наиболее распространенные в Европе), тесно связаны с распространением индоевропейских языков, о чем свидетельствует их присутствие во всех регионах мира, где индоевропейские языки были в древности, от побережья Атлантики в Европе до Иранского нагорья, в том числе почти везде в Европе (за исключением Финляндии, Боснии и Герцеговины), Анатолии, Армении, Южной Сибири, во многих местах в Центральной Азии (в частности, СУАР, Туркменистан, Таджикистан и Афганистан), а также в Иране, Пакистане.
|
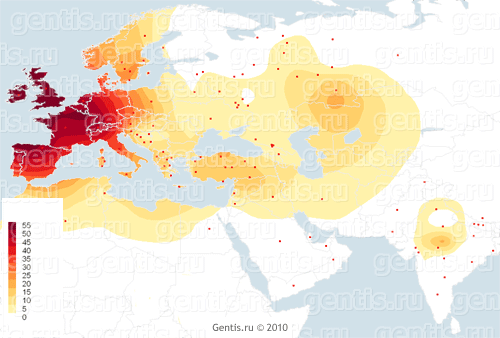
Распределение частот R1b1a2 в современной Европе показывает нам максимальные пики в западной ее части, и постепенное снижение к востоку. Например, в Шотландии, Ирландии и у басков частота линии R1b1a2 достигает 80-90%, а в Испании, Бельгии и Швейцарии 60-70%. Также частоты R1b1a2 ещё очень высоки в северной части Италии и в Германии — порядка 40%. Также европейские субклады более молоды по сравнению со средневосточными или центральноазиатскими. Основная европейская ветвь R-P312/S116 или гаплогруппа R1b1a2 восходит всего лишь к 3500 или 3000 до Р.Х. Это не значит, что старейший общий предок этой линии прибыл в Западную Европу в этот период, но значит, что первый человек, нёсший мутацию R-P312/S116, жил по крайней мере 5000 лет назад, вполне вероятно, где-то в долине нижнего Дуная или в окрестностях Чёрного моря [возможно, как раз в эпоху миграции носителей будущей культуры колоколовидных кубков из Южного Приуралья на Дунай, в Сирию и Иберию]. Захоронения позднего неолита культуры колоколовидных кубков [в Германии] выявили R1b1a2*. |
Гапалогруппа R-U152 имеет удаленный 'восточный' пик частот на севере Башкирии у башкир, где доля этой линии составляет около 72%, в то же время на юге-востоке Башкирии она существенно снижается до примерно 1%. Второй, меньший пик частот - это Центральная Европа [мадьяры].
Гаплогруппу R-M529 можно назвать 'западной атлантической', т.к. мировой максимум частот (94%) зафиксирован на западе Ирландии, в то же время в других регионах Ирландии также максимально высокие значения: на юге (58%), востоке (56%), севере (47%) и юго-западе (45%).
В то время как R1b1 найдена в таком месте, как Левант и Камерун, R1b1b скорее всего возник в Восточной Анатолии.
Ключевые слова для поиска сведений по гаплогруппе R1b Y-ДНК:
На русском языке: Y-хромосомная гаплогруппа R1b, эрбины, R1b-популяция;
На английском языке: Y-haplo-group R1b.
|
|
|
|
|
|