|
|
Главная |
|
|
Пишите |
|
|
Разделы страницы об Y-хромосомной популяции E:
|
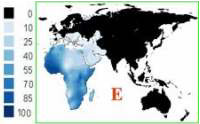
Возраст гаплогруппы E: 52 тыс. лет (45–59 тыс. лет). Происхождение: Восточная Африка (а по версии обратной миграции неоантропов - из Евразии в Африку [кушиты из Албании?]).
SNP-мутации: M40/SRY4064/SRY8299, M96, P29, P150, P152, P154, P155, P156, P162, P168, P169, P170, P171, P172, P173, P174, P175, P176.
Суммарный пик частот приходится на Сомали (77%). Также у арабов Морокко (42%), у эфиопов (20-35%), в Египте (18%), в Судане (17%), в Тунисе (15%). На Ближнем Востоке - в Турции (1-16%), Иране, Ливане и Ираке (10%). В Европе - в Греции (20%), Македонии (17%), Италии (3-16%). На Балканах в среднем встречается с частотой 5-7%. Также как и у дочерней линии E1b1b1a1 по пути на север идет постепенное снижение частот. В России в среднем до 3%. Частично, в невысоких процентах встречается в Азии (0-1%).
Гаплогруппа E1b1b1a1-M78 - самая многочисленная линия в Африке и за его пределами, включая в себя еще 5 крупных ветвей.
|
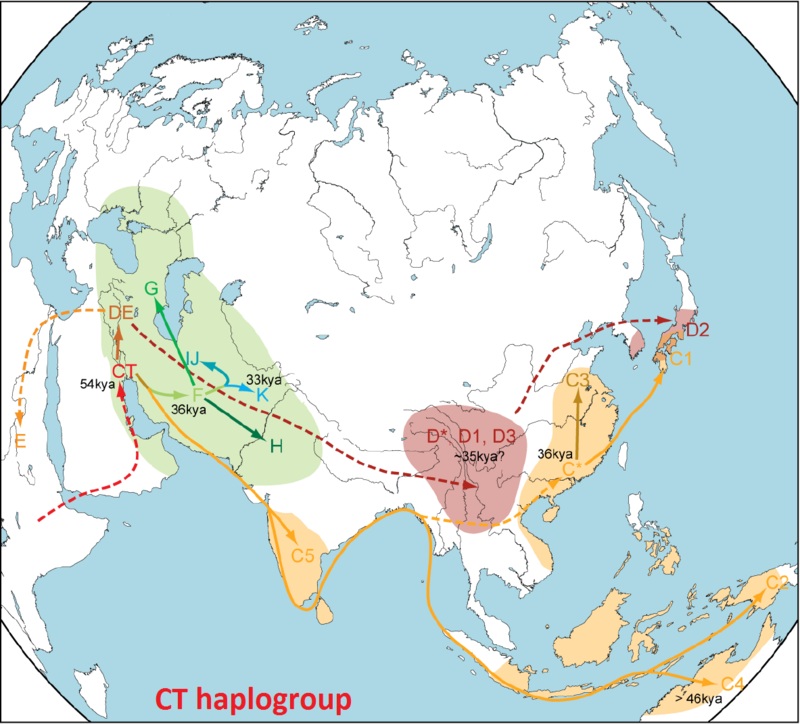
80 т.л.н. Y-хромосомный Адам (или даже 120-250 тысяч лет назад)
|
-----------------
| |
75 т.л.н. A BT
(койс.) |
------------------
| |
70 т.л.н. B CT (миграция через Аравию в Южную Азию)
(пигмеи, хадза) |
70-75 т.л.н. -------------------
| |
65 т.л.н. DE CF
| |
50-55 т.л.н. --------- ---------
| | | |
60 т.л.н. D E C F
(Афр.) (Австр...) (Ар.?)
|
-------------------------
| | | |
55 т.л.н. G IJK H F*
|
|
Согласно гипотезе Семино и др. (2004), гаплогруппа E возникла в Восточной Африке, о чём говорит плотность концентрации её субкладов на территории Африки сегодня. Гаплогруппы E1a и E2 встречаются почти исключительно в Африке, и только лишь E1b1b1 с высокой частотой встречается в Европе и Западной Азии, наряду с Африкой. Большинство африканцев субсахарской области относятся к иным подклассам гаплогруппы, чем E1b1b1, тогда как большинство неафриканских носителей относятся как раз к подклассу E1b1b1.
|
Род E1b1b1 (snp М35) объединяет около 5% всех мужчин на Земле и насчитывает около 700 поколений до общего предка. Родоначальник рода E1b1b1 родился примерно 15-20 тысяч лет назад на Африканском Роге (возможно, в Северной Эфиопии) или на Среднем Востоке (возможно, в Йемене). По расовой принадлежности хамиты относились к Кушитской большой ветви Западного расового ствола. Согласно теории Дьяконова-Бендера, в Эфиопии прахамито-семитский язык выделился из праностратического языка примерно 14 тыс. лет назад.
Примерно 13 тыс. лет назад племена, принадлежащие этому роду, начали своё расселение на север и на юг от Эфиопии. Около 10 тыс. лет назад род E1b1b1 распространился в районе Средиземноморья и в Южной Африке.
Северные ветви рода E1b1b1:
Южные ветви рода E1b1b1:
Среди русских представлена только одна ветвь - E1b1b1a2 (V13), идущая от переселенцев с Балкан.
Потомки северных ветвей рода E1b1b1b живут сейчас, в основном, в Северной Африке, Западной Азии, Западной и Восточной Европе. Среди них - актер и певец Андриано Челентано.
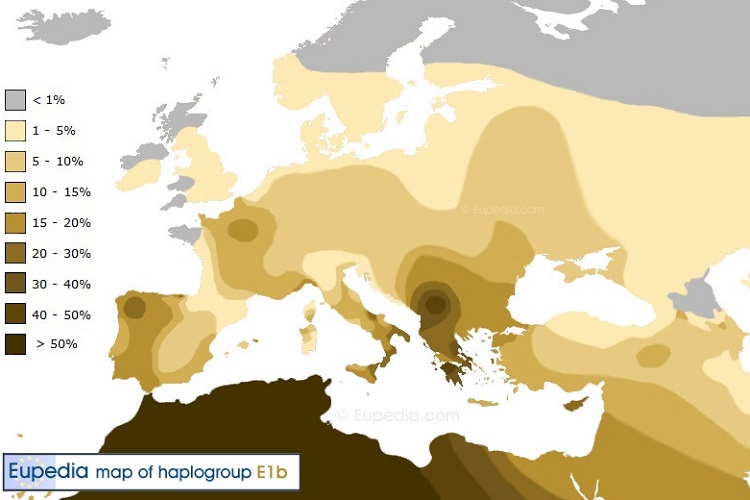
Гаплогруппа E1b1b1 (ранее E3b) представляет последний крупный поток миграции из Африки в Европу. Считается, что впервые E1b1b1 появились в районе Африканского Рога и Южной Африки примерно 26 000 лет назад и разошлись на Ближнем Востоке в эпоху мезолита. На европейском континенте они имеет самую высокую концентрацию на северо-западе Греции, Албании и Косово, а затем она уменьшается при удалении от Балкан в остальной Греции и Западной Турции. За пределами Европы, также в большинстве стран Ближнего Востока, Северной и Восточной Африке, особенно в Марокко, Ливии, Египте, Йемене, Сомали, Эфиопии и Южной Африке гаплогруппа E1b1b1 имеет очень высокое распространение.
Разве E1b1b1 могли переселиться прямо из Северной Африки в Европу в связи с изменением климата? До сих пор неясно, когда гаплогруппа E появилась в Европе. Последние тесты ДНК из неолитических стоянок в южной Германии и южной Франции не показывали след E1b1b1. Это свидетельствует о более позднем прибытии, либо к концу неолита / энеолита, либо в эпоху бронзы. В случае отсутствия Y-ДНК из неолита Греции, Южной Италии и Иберии, ничего не исключает возможности того, что гаплогруппа E1b1b1 присутствовала в этих регионах с неолита, мезолита или даже позднего палеолита.
В Северной Африке носители E1b1b1 пересекли Средиземное море (вероятно, несколькими независимыми волнами миграций) в какое-то время с последнего ледникового максимума (около 20 000 лет назад), или последнего опустынивания Сахары, который начался, когда муссон отступил на юг 6 200 лет назад. В последний ледниковый максимум (20 т.л.н.), уровень моря был на 120 метров ниже, чем сегодня, и Гибралтарский пролив был шириной всего несколько километров, что позволяло использовать даже самые примитивные плоты, чтобы легко его пересечь.
Совпадение ли, что последние следы неандертальцев в Иберии датируются 24 000 лет назад? Может ли их исчезновение являться результатом поглощения кроманьонцами из Северной Африки? Последние неандертальцы иберийского п-ва обнаруживают некоторые признаки гибридизации с Homo Sapiens. Бесспорно, что Homo Sapiens колонизировали Европу из Ближнего Востока, однако, вполне вероятно, были и колонизации с северо-запада Африки, приводящие к противоборству между двумя видами людей. Это объясняет, почему существует так много Северо-Западно африканских E-M81 в Португалии и северо-западной Испании, которая не подтверждается ни исторически засвидетельствованными миграциями, ни археологически очевидными неолитическими миграциями из Северо-Западной Африки.
Пустыня Сахара неоднократно меняла свой ландшафт от пышных зеленых джунглей до горячих и засушливых пустынь за последние 20000 лет. Наступление более теплого и засушливого климата в конце ледникового периода 13000 лет назад вновь привело к появлению тропических муссонов с 10000-7000 лет до настоящего времени. Опустынивание, происходящее сегодня началось около 4200 гг. до нашей эры. Эти серьезные преобразования окружающей среды, безусловно были сильным воздействием на коренные линии E1b1b, в результате чего население в эпоху неолита мигрировало из Сахары. Также неудивительно, что люди из Северной Африки пересекали Средиземное море и в конце неолита. Области, наиболее пострадавшие от опустынивания должны были быть в районе современной Ливии. Северный Магриб находится под защитой гор, которые остановливали наступление пустыни. [Также опустынивания не было в долине Нила и вокруг его дельты в Египте]. Согласно одной из гипотез неолитическое население Ливии мигрировало в современные Южную Италию, Грецию, Македонию и Албанию, принеся с собой линии E-V13, которая в настоящее время все еще находится в Ливии, а также в Иберии, Египте и Леванте, хотя гораздо плотнее вокруг Греции. Кроме того, вместо пересечения непосредственно Средиземного моря из Туниса до Сицилии, затем в Италию и на юг Балкан, миграции могли доходить до Греции по средиземноморскому побережью через Египет, Левант и Анатолию. Некоторые мигранты могли идти по опбережью на запад к Иберии, что объясняет, почему E-V13 имеется в западной Иберии, наряду с магрибскими E-M81, в то время как греки никогда не колонизировали этот регион.
Гаплогруппа E1b1b1a (или E-M78, ранее E3b1a) является наиболее распространенным среди различных субкладов гаплогруппы E среди европейцев и на Ближнем Востоке. E-M78, как полагают, мигрировали из Египта в эпоху мезолита и неолита до колонизации Ближнего Востока, где она смешивается с коренными жителями принадлежащими гаплогруппам J и G. Финикийцы, из Леванта, также способствовали распространению E1b1b1a (равно как и J2, Q и T) в таких странах, как Кипр, Мальта, Сардиния, Ибица и южная Иберия. Низкий уровень E1b1b1a в Сирии и Анатолии почти наверняка связан с конкуренцией со стороны других основных гаплогрупп неолита: G2 и J2.
Субклад E-M78 делится на 4 основных ветви: E1b1b1a1 (E-V12) E1b1b1a2 (E-V13) E1b1b1a3 (E-V22) и E1b1b1a4 (E-V65), каждая в свою очередь подразделяются на "а" и "б" подсубклады.
Гаплогруппа E1b1b1a1a-V12 более всего найдена у арабов южного Египта (44%), у арабов из Бахарии (15%), у арабов египта близ Луксора (9%), у арабов дельты Нила (6%). Также встречается в Турции с частотой около 4%.
Субклад E-V12 является самым распространенным из субкладов M78 в Египте. Его низкое присутствие в Греции и Анатолии, показывают, что он, вероятно, уже существовал, когда E переехали туда в начале неолита.
Гаплогруппа E1b1b1a1b-V13 - самая распространенная на данный момент линия за пределами Африки. Встречается преимущественно на Балканах, а также частично на Ближнем Востоке. Достигает пика в Греции (17%), Албании (32%), Болгарии (16%). По пути на север идет постепенное снижение частот этой линии. В Италии (5-8%), в Украине и Венгрии (9%), Словакии (8%), в Австрии, Чехии, Португалии и Германии (4-5%), в России (1-3%). В северной Африке (0-3%), на Б. Востоке (1-5%).
Субклад E-V13 является одним из основных маркеров неолита и сельскохозяйственных миграций из Леванта. Как и все другие субклады из E1b1b1a, E-V13 возник в Северо-Восточной Африке примерно в конце последнего ледникового периода. Его частота составляет в настоящее время гораздо выше, в Греции, Южной Италии и на Балканах, чем где-либо в связи с эффектом основателя, т.е. миграции были у небольшой группы поселенцев в основном этой линии (а также небольшое количество других Северо-Восточной африканских линий, в частности E-M123 и T). Археологические данные показывают, что область Фессалии, на севере Греции, была отправной точкой (ок. 6000 до н.э.) для распространения сельского хозяйства через Балканы и бассейн Дуная, в северной Франции на западе и Россией на востоке. Современное распределение E-V13 намекает на сильную корреляцию с неолитом и энеолитом Старой Европы, такими как Культура Винча, Боян, Марица и Караново.
Тем не менее, генетические тесты трех мужчин образцы культуры LBK только показал наличие гаплогруппы F и G2a. Пример, очевидно, слишком малы, чтобы исключить, что E1b1b также попал в Европу в эпоху неолита. E-V13 также связано с древнегреческим распространением и колонизацией. За пределами Балкан и Центральной Европы особенно часто встречается в Южной Италии, на Кипре и Южной Франции - всех частях классического древнегреческого мира [древние греки даже в Южной Франции плотно жили?].
Гаплогруппа E1b1b1a1c-V22 встречается в Африке у эфиопов (25%), у арабов из центрального Египта (22%), а также у арабов из дельты Нила (14%) и у морокканцев (8%). Кроме Африки - в Италии (1-4%), Австрии (4%), Испании (3%), Македонии и Греции, а также в Палестине (6-9%).
Субклад E-V22 является преобладающим субклад в Леванте и, следовательно, связан с финикийцами и евреями, в дополнение к распространению земледелия. Финикийцы могли бы распространить E-V22 на Сицилии, Сардинии, на юге Испании и Магриба, а евреи в Грецию и на материке Италии и Испании. Тем не менее, средиземноморский маршрут для распространения сельского хозяйства проходил через те же регионы. Поэтому невозможно узнать, в настоящее время, какой из двух периодов (неолита или античности) сыграл значительную роль в распространении V22 по всему Средиземноморью.
Гаплогруппа E1b1b1a1d-V65 распространена в основном у берберов Морокко (30%), ливийских арабов (20%), египетских берберов, а также частично в Европе у итальянцев (Сицилия, Сардиния).
Субклад E-V65 находится в Северной Африке, с максимальной частотой в Ливии и Марокко. Кроме того, вероятно, возник в Египте. В Европе она находится на низких частотах в Греции и Сицилии, но интересно, что составляет одну четвертую сардинского E. Это может быть связано с миграцией из финикийских колоний в странах Магриба и Сардинии (в Сардинии гаплогруппа I2a1 также присутствует на низких частотах, а также на побережье Алжира и Туниса, подтверждающие обмен населением между двумя регионами, может быть, когда обе территории были финикийскими колониями).
Гаплогруппа E1b1b1a1e-M521 - очень редкая ветвь, которая найдена только в Греции в небольших количествах (2%).
Гаплогруппа E1b1b1b (E-M81, ранее E3b1b) характерно для берберов Северо-Западной Африки. В некоторых районах Марокко E1b1b1b может достичь своего пика в 80% населения. Этот субклад также находится в Иберии, Италии и южной Франции, с наибольшей концентрацией на юге Португалии (12%) и снижается по мере продвижения на север.
Гаплогруппа E1b1b1b1-M81 характерна для берберов северо-западной Африке. В некоторых районах Морокко достигает частоты 80%. Найдена также в Иберии, Италии и на юге Франции. Наиболее сконцентрирована в южной части Португалии (12%) с уменьшением частоты по пути на север.
Гаплогруппа E1b1b1c (E-M123) и ее основные ветви E1b1b1c1 (E-M34) также связаны с распространением сельского хозяйства и древними ближневосточными цивилизациями. Эта гаплогруппа достигает пика в южном Леванте (10-12% в Палестине и Ливане), откуда он расширяется во все стороны на Ближнем Востоке, Северной Африке, Южной Азии и Юго-Восточной Европы. Распределение E-M123 почти полностью совпадает с расширением сельского хозяйства в период неолита. E-M123, кажется, идут рука об руку с гаплогруппой G2a, с той лишь разницей, что G2a достигает максимальной частоты примерно на Кавказе и в Анатолии, где крупный рогатый скот, свиньи и козы были впервые одомашнены. Внутри Европы, E-M123 более или менее распространен субклад E-V13, с высокой частотой (от 1 до 5%) наблюдается в Греции, Южной Италии, на Балканах и в бассейне Дуная, но затухание по отношению к Германии, Польши, Украине и России, где его частота составляет менее 1%.
Гаплогруппа E1b1b1c-M123 распространена в Европе в основном на юге Италии (13%), на Балканах (1-4%). На Ближнем Востоке - в Омане (11%), в Турции (3-9%), в Ливане (4%). На севере Африки в Тунисе (5%), в Алжире (3%), а также в Эфиопии (2-5%).
Гаплогруппа E1b1b1d-V6 встречается среди этнических групп Эфиопии (14%) и Сомали.
Гаплогруппа E1b1b1e-M293 найдена в этнических группах Танзании (датог, кхве, сандаве, бурунге и т.д.) и Южной Африки.
Гаплогруппа E1b1b2-M281 найдена в небольших количествах в Ефиопии.
Y-хромосомная популяция E2 (как и E1a) встречаются почти исключительно в Африке.
Ключевые слова для поиска сведений по Гаплогруппе E Y-ДНК:
На русском языке: Y-хромосомная гаплогруппа E, человеческая популяция мужской E-гаплогруппы, популяционная группа E Y-ДНК человека;
На английском языке: Y-haplo-group E.
|
|
|
|
|
|